ty, located at the distal end of the anti-back surface of the bones of the metacarpus, metatarsus and distal phalanges of the fingers (see skeleton). The sesamoid bones include the patella and the accessory bone of the wrist.
BRIEF INFORMATION ON PHYLO AND ONTOGENESIS OF MUSCLES
Phylogenetic transformations. Muscular elements in a row
Orbits of living beings appear early in coelenterates. They are not yet separated into independent morphological units, but are only contractile muscle elements of epithelial cells. In the future, they are isolated from the epithelium, forming several layers of smooth muscle cells, closely connected with the skin, resulting in the formation of the so-called musculoskeletal sac (flatworms). The source of the formation of muscle cells is the mesoderm.
With the appearance of a secondary body cavity, the musculature is divided into somatic, which is part of musculocutaneous sac, and visceral, surrounding the intestines and blood vessels. Despite this division, it can be either all smooth (annelids) or all striated (insects). This indicates that in phylogenesis, striated muscles almost do not differ from smooth muscles either in origin or in function. With further complication of organization, somatic and visceral muscles develop divergently, more and more diverging from each other structurally and functionally.
At In primitive chordates (lancelet, cyclostomes), all somatic muscles develop from mesoderm somites and are striated. It is a pair of right and left longitudinal muscles running along the entire body, divided by connective tissue septa - myoseptami into a number of myomers - short segments from straight muscle bundles. Such (segmental) division of a single muscle layer is called metamerism (Fig. 73).
With the isolation of the head and the development of the limbs (in the form of fins) differentiate the musculature. The longitudinal muscle in fish is divided by a horizontal septum into dorsal and ventral muscle. They are innervated by the dorsal and ventral branches of the spinal nerves, respectively. This innervation is preserved during all further transformations of the muscles. The dorsal and ventral longitudinal muscles, due to the uniformity of movements of primary aquatic animals, have a myomeric structure. Each myomer usually corresponds to its own vertebra and a paired spinal nerve. In higher fishes (herrings and others), one can see their longitudinal splitting into separate layers. The musculature of the fins is also isolated, however, in comparison with the muscles of the body trunk, it is poorly developed, since the main load during movement in aquatic animals falls on the tail and trunk.
Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
Rice. 73. Muscles of the body of chordates:
A - lancelet; 5 - fish; B - tailed amphibian; G - reptile; 1 - myomers (myotomes); 2- myosepts; 3- dorsal m. of the body; 4- longitudinal side partition; 5 - dorsal m of the tail; 6 - surface compressor; 7- trapezoidal m.; 8 - ventral m. of the body; 9 - ventral m. tail; 10 - mm. chest limb; 11 - widest m, back; 12, 13, 14 - ventral mm. (12 - oblique external, 13 - oblique internal, 14 - straight); 15 - mm. pelvic limb.
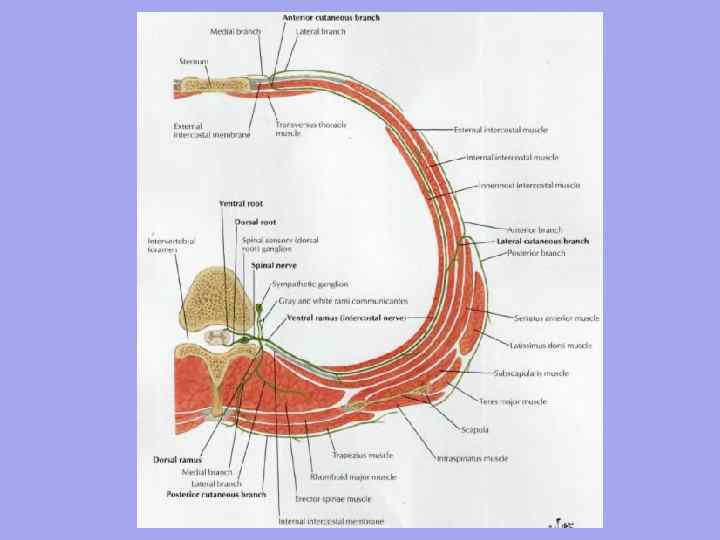
With access to land and an increase in the variety of movements, the division of muscle layers into separate muscles grows both along and across. In this case, metamerism gradually disappears. It is well traced in the muscles of fish, is also noticeable in amphibians, weakly in reptiles. In mammals, it is preserved only in the deep layers, where the short muscles connect the elements of two adjacent bone segments (interspinous, intertransverse, intercostal muscles) to each other.
Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
First of all, metamerism begins to disappear in the abdominal part of the body, where, already in amphibians, individual myomeres merge with the formation of wide lamellar abdominal muscles. Along with this, there is a longitudinal splitting of the muscular abdominal wall with the formation of a four-layer abdominal press. In the dorsal muscles of the body of amphibians, two strands can be distinguished: lateral and medial, the metamerism of which is obscured only in cervical region where independent muscles are isolated.
At reptiles, the muscle bundles of the lateral and medial muscle cords acquire a different direction. Myomeria is preserved only in the deep layers. The closer to the head, the clearer the fragmentation of the dorsal strands into separate muscles.
At mammals, the somatic musculature is differentiated to the greatest extent. In the dorsal muscles, 4 layers are formed due to the stratification of the lateral and medial muscle cords. At the same time, a clear pattern is observed: the deeper the muscle is, the better its metamerism is expressed; the closer to the outer surface of the body the muscle lies, the more it loses metamerism, spreading through the body in a wide layer. The dissection of the dorsal muscles also increases in the cranial direction, which is associated with the degree of mobility of the spine. If in the region of the sacrum - the most immobile part of the stem skeleton
- the dorsal muscles are absolutely not dissected, then in the area of the withers, and especially the neck, the muscle complexes consist of a large number of independent muscles.
The ventral musculature of the stem part of the body also has 4 layers, although not fully expressed everywhere. In the chest, these are the internal and external intercostal, rectus and transverse pectoral muscles, in the lumbar-abdominal region - the abdominal muscles.
The locomotor function of the muscles of the tail becomes less and less with landfall and is completely lost in mammals. This leads to a significant decrease in muscle mass while maintaining a high degree of their differentiation due to the mobility of the tail.
The limbs of terrestrial vertebrates originated from a lobe-fin, very mobile, with a well-developed skeleton and strong muscles (coelacanth). The metamerism of the muscles of the limbs, which is clearly visible in ray-finned fishes, is lost very early in phylogenesis, especially with landfall. With the transformation of a limb into a complex lever that supports and moves the animal's body on land, a large number of muscles become isolated.
For primitive tetrapods, the humerus and femur bones are characteristically moving away and up from the belt. With such an arrangement of the limbs, large expenditures of muscle energy are required to maintain the body in a hanging position. On the thoracic limb, the greatest load falls on the coracoid bone, to which, as a result, the bulk of the muscles of the shoulder and elbow joints are attached.
Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
Adaptations for fast running, manipulation of the thoracic limb, and the ability to rest while standing, developed in mammals, were accompanied by rotation of the limb from the segmental to the sagittal plane, opening of the joints, and ever higher elevation of the body above the ground. At the same time, the conditions for the action of gravity and the work of muscles during the standing and movement of the animal changed. In ungulates, the adaptation of the limbs to rapid forward movement and to the economical expenditure of muscle energy when standing has led to the loss of variety of movement. This was expressed in an even greater reduction of the shoulder girdle (disappearance of the clavicle) and straightening of the free limb. The shoulder girdle lost its bone connection with the axial part of the body and acquired an extensive support area with the help of muscles that connected it with the head, neck, withers, back and chest. So the musculature of the limbs in terms of mass began to prevail over the musculature of the body. The muscles of the girdle and proximal parts of the limbs largely cover the top of the trunk muscles and partially displace it. The development of the muscles of the distal links is largely determined by the peculiarities of the mechanics of movement and the ecology of the animal (walking, crawling, jumping, digging, etc.). In ungulates, in connection with the reduction of fingers, straightening of the joints, there was a decrease in the number and complication of the structure of the muscles of the distal parts of the limbs.
And finally, the most superficial and least dissected muscle layer is the subcutaneous muscles - part of the somatic muscles that first appeared in reptiles. In mammals, it is highly developed, especially in animals that can coagulate (hedgehog, armadillo). Of domestic animals, it is well developed in the horse and has the appearance of wide layers lying under the skin in the neck, withers, shoulder blades, chest and belly (see Fig. 72). On the head, the subcutaneous muscles come into close contact with the visceral and enter integral part in the muscles of the face, eyelids, nose, auricle.
Complex transformations in the musculature of the head occur in parallel with complex phylogenetic transformations of the skull. As a result, the somatic muscles in the head area are largely replaced by the visceral muscles surrounding the head gut. The somatic musculature of the head is already in fish, represented only by the muscles of the eye and some supra- and subgill muscles with a longitudinal direction of the muscle fibers (they participate in the respiratory movements of the gill apparatus).
The visceral muscles surrounding the head end of the intestinal tube underwent significant differentiation, acquired the properties of striated muscle tissue, but retained its annular direction of fibers. It formed circular muscle layers of the jaw, hyoid and gill arches, on the basis of which the bulk of the head muscles develop: jaw, hyoid, gill, some muscles of the shoulder girdle with grasping, chewing and other functions.

Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
In mammals, the somatic muscles of the head are represented by the muscles of the eye, middle ear, tongue, and some muscles of the hyoid bone. Visceral muscles form mimic (facial) and chewing (jaw) muscles.
And finally, only mammals have a muscular abdominal barrier - the diaphragm.
ontogenetic development. The somatic musculature mainly comes from the myotomes of the somites of the mesoderm (Fig. 74). In the region of the head, the muscles of the eyeball are formed from the three anterior myotomes. The anterior behind-the-ear myotomes disappear, and the hyoid muscles develop from the posterior (occipital) ones. Visceral muscles of the head of mesenchymal origin. Cervical, thoracic, lumbar, sacral and caudal myotomes are laid in accordance with the number of metameric body segments. They grow in the dorsal and ventral directions and give rise to all the somatic muscles of the neck, trunk and tail. The muscles of the extremities are formed by outgrowths of the ventral sections of the myotomes, to which the cellular material is attached, which is expelled from the parietal sheet of the mesoderm splanchnotome. The anlage of the musculature somewhat lags behind the anlage of the skeleton and to a certain extent depends on it.
Rice. 74. Metameric anlage of muscles in the embryo of a mammalian myotoma:
1 - occipital. 2 - cervical, 3 - chest. 4 - lumbar, 5 - sacral, 6 - tail.
In the embryonic period, from the 20th-22nd day of development, in a large cattle in myotomes, myoblasts multiply. In the prefetal period, anatomical differentiation begins: muscles and muscle groups are isolated. In parallel with this, but much longer is the histogenesis of muscle tissue. Myoblasts merge into muscle tubules, myofibrils appear in them. Anatomical differentiation basically ends in the prefetal period - by the 50-55th day. The laying and differentiation of muscles goes in a certain sequence. Before others, the axial muscles are laid. In it, differentiation proceeds from the head end to the tail. At the same time, deep muscles differentiate earlier
Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
superficial. In the process of muscle differentiation, the corresponding cranial or spinal nerves grow into them. This connection is established very early and persists throughout life. Bookmarks of the limbs appear in the form of roller-like thickenings near the ventral areas from the 5th cervical to the 1st thoracic myotome - the rudiment of the thoracic limb and from the 1st lumbar to the 3rd sacral myotome - the rudiment of the pelvic limb. Soon, the rollers are pulled together and take the form of flattened conical outgrowths - kidneys. The formation of muscles on the thoracic limb in the calf embryo begins from the 32nd day, and on the back - from the 34th day embryonic development. Before others, the muscles of the belts are laid, then the free limb, where the process spreads from the proximal to the distal links. As in the axial part of the body, differentiation of the deep muscles occurs earlier, and that of the superficial ones later. On the lateral side of the limb, extensors, abductors and arch supports are laid, on the medial side - flexors, adductors and pronators. The abdomens of the muscles are laid before the tendons. By the end of the prefetal period, the muscles of the limbs are anatomically formed, but histologically immature - they consist of muscle tubes lying in bundles. During the fetal period, the histological differentiation of muscles continues: the number and size of muscle tubes increase, the tubes turn into muscle fibers, the number of myofibrils increases in them; endomysium and perimysium of muscles are formed, capillary networks develop, bundles of I, II and III orders are formed.
As a result of anatomical and histological differentiation, the dorsal muscles of the spinal column are formed from the dorsal sections of the myotomes, lying above the vertebral bodies. It is innervated by the dorsal branches of the spinal nerves. From the ventral sections of the myotomes, the ventral musculature of the spinal column is formed, which lies under the bodies of the vertebrae, the musculature chest, abdominal wall and diaphragm. From the muscle kidneys, all the muscles of the limbs develop.
AT In the process of organogenesis, the muscles separate along the length, thickness, crushing or fusion, the formation of complex and multifid muscles, the formation of their pinnate structure. In the early fetal period, the muscles of the trunk grow faster, and in the late period, the muscles of the limbs, especially their most distal links - the paws.
In ungulates, by birth, the apparatus of movement is fully formed, which immediately begins to function: after a few hours, a newborn calf, lamb, foal, piglet can follow the mother. However, this does not mean that the processes of growth and differentiation have been completed in the locomotor apparatus. They continue until the age of morphophysiological maturity, and the adaptive restructuring of the movement apparatus occurs throughout life.
postnatal muscle growth. After birth, intensive growth of muscles continues, which overtakes the skeleton in terms of growth rate. This process is especially intense in the first two months after birth.
Vrakin V.F., Sidorova M.V. |
MORPHOLOGY OF FARM ANIMALS |
Denia. The next growth peaks in cattle occur at the 6th and 12th months of life, in sheep - at the 3rd and 9th months. The axial muscles grow faster than the muscles of the limbs, especially with the onset of puberty. In newborn calves, the mass of axial muscles is 46%. and in 14-month-olds - 53%. On the extremities, there is a high rate of muscle growth in the proximal links (compared to the distal ones). On the thoracic limb, they grow somewhat more intensively, but complete their growth faster than the muscles of the pelvic limb. The extensors grow faster than the flexors, and the periods of rise in their growth rate do not coincide.
With age, the number of muscle fibers per unit area in the muscles also decreases in the primary muscle bundles, since along with the thickening of the muscle fibers (about 15-20 times), the muscles grow into the connective tissue, it becomes denser, the muscle bundles. I order include fewer fibers. However, the relative amount of connective tissue in muscle decreases with age, and muscle
Increasing. So, for 18 months in bulls, the amount of connective tissue increases by 8 times, and muscle - by 17 times. The chemical composition also changes: the amount of protein and fat increases, water becomes less. Each type of muscle has its own dynamics of chemical parameters.
Not only muscle groups, but each muscle has its own character of growth, which is associated both with the peculiarities of its internal structure and functioning. The highest growth rates in muscles of the dynamic type. The uneven growth of muscles largely determines the change in the proportions and articles of the body.
Influence of internal and external factors on muscle growth. The way of life of the animal, the method of production and the nature of the food leave an imprint on the growth and differentiation of the muscles. So, pigs develop more dorsal muscles, especially necks. The horse has better developed chewing muscles than cattle. The abdominal muscles, on the contrary, are more developed in cattle.
The gender of the animal also affects the nature of muscle growth. With the same fatness in bulls, the muscles are better developed and make up a larger percentage in the carcass than in heifers and castrated bulls. In addition, the growth of muscles in bulls lasts longer, therefore, more meat can be obtained from them in the end. In bulls, the muscles of the neck, withers and shoulder girdle are more developed (which is important for the strength of the animal when establishing a hierarchy in the herd). In heifers, the muscles of the abdominal press and the back half of the body are more developed. Castrates by the nature of muscle growth approach heifers, and by the growth of the longest and semispinous muscles they lag behind animals of both sexes. Gobies have fewer fatty inclusions in their muscles, while heifers and castrates have thinner muscle fibers, and marbling of meat is well expressed.
There are also some differences in the rate of growth and development of muscles between breeds with different directions of productivity. Early maturing breeds are characterized by high growth energy, but late maturing
The complex phylogenetic development of the muscles of the trunk, neck and head is due to the fact that they originate from two rudiments: one part is the body's own muscles, the other is laid in the lateral plates related to the intestinal muscles associated with the mesenchyme of the gill apparatus.
For a more correct representation of the evolution of muscles, we will analyze their rudiments.
The phylogeny of the muscles of the trunk. Lower vertebrates, like the lancelet, have paired muscles located on the sides of the body. Lateral muscles with the help of a horizontal lateral connective tissue septum are divided into dorsal and abdominal sections (Fig. 186). Each myomer is separated from the neighboring one by a vertical connective tissue septum (myoseptum); these partitions are located transversely to the body. The vertical septa in the area of the lateral horizontal connective tissue septa are bent and form an angle facing forward. The upper (dorsal) and lower (ventral) ends of the myomeres also curve forward. It turns out a broken line between the myotomes, similar to such a figure, where the angle located on the lateral connective tissue groove is facing forward. In the form of such cones, all the dorsal muscles of the lower vertebrates are built. But already in some fish, differentiation is noted in the position and direction of the myotomes. In the lateral wall of the abdomen, part of the myotomes is located obliquely to the sagittal plane at different depths. This shows that in aquatic animals the first signs of restructuring of myotomes appear, which in terrestrial vertebrates turn into the lateral oblique muscles of the abdomen. In the middle of the abdomen, part of the myotomes is transformed into the rectus abdominis muscle.
The dorsal musculature of terrestrial vertebrates is adapted to the displacement of individual body segments, since the myotomes are thrown over those joints where the movement takes place. In amphibians, the back muscles are built from independent myotomes and resemble the muscles of lower vertebrates. Only in reptiles is the disappearance of the horizontal connective tissue septa and the blurring of the boundary between the dorsal and ventral muscles. Due to the increased mobility of the spine and chest, some of the myotomes merge into larger muscles. This is how mm. interspinales, intertransversales, transversospinales, longissimus, iliocostalis and occipital-vertebral group. This led to the fact that in higher vertebrates, including humans, primary myotomes remained only in the form of short muscles mm. rotatores, connecting successively segments of the body (vertebrae).
186. Superficial abdominal muscles of a newt (according to Maurer).
1 - subgill muscle: 2 - intermuscular septum; 3 - m. obliquus externus superficialis; 4 - m. rectus interims; 5 - m. rectus superficialis.
The abdominal muscles in lower amphibians still retain division in the form of separate myotomes (Fig. 186), but in the middle there are longitudinal myotomes fused into the rectus abdominis muscle. In the side walls, the myotomes change direction, but do not yet form independent muscle layers. In higher vertebrates, the myoseptae disappear and the myotomes merge with each other into large muscle layers arranged in three layers. These muscle layers in humans are represented by three lateral muscles. Only in the rectus abdominis muscle, in all animals, primary segmentation is preserved in the form of tendon bridges intersectiones tendineae.
AT thoracic region ribs grow along the connective tissue septa between the myotomes, and intercostal muscles are located in the intercostal spaces, representing the continuation of the abdominal myotomes. From the ventral side of marsupials m. pyramidalis reaches the sternum. In mammals, this muscle is preserved as a rudiment.
Phylogeny of the visceral muscles. In lower vertebrates, the anterior intestine is formed by circular fibers covering the entire visceral apparatus in the form of a common constrictor. Beginning with cyclostomes and selachians, deep muscle bundles come into contact with visceral arches, which are covered from the outside by lateral muscles perforated by gill openings. Separation of individual muscles is observed in the muscle mass: a muscle is attached to the upper section of the I branchial arch, which lifts the palatine-square cartilage (innervated by the V pair of cranial nerves), to the lower section is a muscle that leads the lower part of the II branchial arch (innervated by the V pair of cranial nerves), intermaxillary muscle, lying between the branches of the lower jaw (innervated by the V and VII pairs of cranial nerves). From the side of the back behind the gill slits rear end common decompressor is isolated in m. trapezius.
Beginning with amphibians, in connection with the fixation of the upper jaw to the skull, the muscles of the visceral apparatus are transformed. The muscle that lifts the palatine-square cartilage in amphibians is transformed into a muscle that raises the eye, which is reduced in higher animals. From the muscle that leads the lower jaw, chewing muscles are formed.
The intermaxillary muscle in amphibians and reptiles retains its position, and in higher animals it turns into the chin-hyoid muscle. A bundle is separated from the intermaxillary muscle to form the anterior belly of the digastric muscle. All these muscles are innervated by the fifth pair of cranial nerves.
Aquatic animals have muscles that are innervated by the VII cranial nerve. This includes the muscle that lowers the lower jaw, which in mammals turns into the posterior belly of the digastric muscle (innervated by the VII pair of cranial nerves).
In reptiles, the neck constrictor reaches a significant development (innervated by the VII cranial nerve), which in mammals is divided into superficial and deep parts. The muscles of the perioral fissure develop from the deep part, and the rest of the mimic muscles are formed from the superficial part (it is innervated by the VII pair of cranial nerves).
The muscles associated with the gill apparatus, with the loss of the gill type of breathing, are converted into the muscles of the larynx, pharynx and hyoid. The trapezius muscle loses its connection with the gill arches and passes to the shoulder girdle. The sternocleidomastoid muscle splits off from its anterior edge.
In fish and terrestrial vertebrates, the hyoid muscles develop from the abdominal processes of the occipital myotomes, which go around the gill apparatus behind and are located on the ventral side under the visceral apparatus. From the rectus abdominis in amphibians and other more highly organized animals, the muscles of the tongue, the hyoid, and the muscles lying below the hyoid bone develop.
LECTURE MYOLOGY PHYLOGENESIS, ONTOGENESIS AND FUNCTIONAL ANATOMY OF THE MUSCLE SYSTEM Performed by: Vladimirova Ya. B. Kokoreva T. V.

 Muscles or muscles (from Latin musculus - mouse, small mouse) - organs of the body of animals and humans, consisting of elastic, elastic muscle tissue that can contract under the influence of nerve impulses. Designed to perform various actions: body movements, contraction of the vocal cords, breathing. Muscles are 86.3% water. There are 640 muscles in the human body
Muscles or muscles (from Latin musculus - mouse, small mouse) - organs of the body of animals and humans, consisting of elastic, elastic muscle tissue that can contract under the influence of nerve impulses. Designed to perform various actions: body movements, contraction of the vocal cords, breathing. Muscles are 86.3% water. There are 640 muscles in the human body
 Motivation: - - - the possibilities of the movement being made, the volume of movement; active or passive movements are triggered by one or another muscle group; acting on the muscular apparatus, we change the general state; muscle relief is a guide for the topography of blood vessels and nerves; muscle transplantation, that is, the muscle can be "retrained".
Motivation: - - - the possibilities of the movement being made, the volume of movement; active or passive movements are triggered by one or another muscle group; acting on the muscular apparatus, we change the general state; muscle relief is a guide for the topography of blood vessels and nerves; muscle transplantation, that is, the muscle can be "retrained".
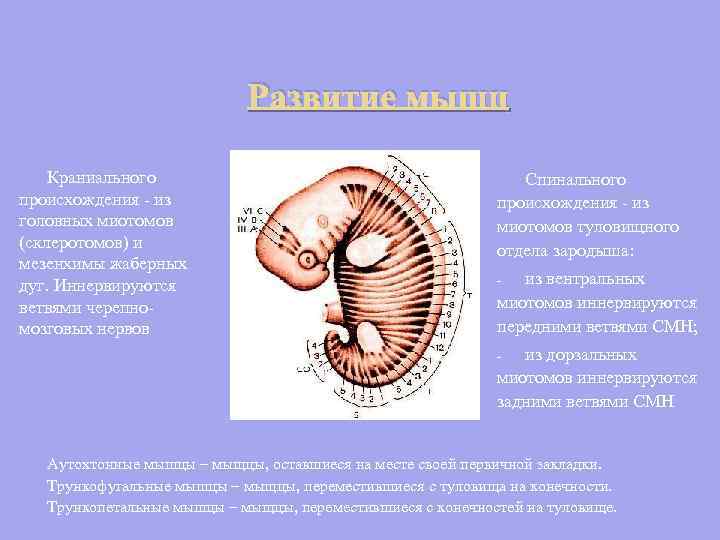 The development of muscles of cranial origin - from the head myotomes (sclerotomes) and the mesenchyme of the gill arches. Innervated by branches of the cranial nerves Spinal origin - from the myotomes of the trunk of the embryo: from the ventral myotomes are innervated by the anterior branches of the SMN; - from the dorsal myotomes are innervated by the posterior branches of the SMN - Autochthonous muscles - muscles that have remained in the place of their primary laying. Truncofugal muscles are muscles that have moved from the trunk to the limbs. Truncopetal muscles - muscles that have moved from the limbs to the trunk.
The development of muscles of cranial origin - from the head myotomes (sclerotomes) and the mesenchyme of the gill arches. Innervated by branches of the cranial nerves Spinal origin - from the myotomes of the trunk of the embryo: from the ventral myotomes are innervated by the anterior branches of the SMN; - from the dorsal myotomes are innervated by the posterior branches of the SMN - Autochthonous muscles - muscles that have remained in the place of their primary laying. Truncofugal muscles are muscles that have moved from the trunk to the limbs. Truncopetal muscles - muscles that have moved from the limbs to the trunk.

 Striated Smooth 1. The unit of organization is the myocyte. The length is about 50 µm. Width from 6 µm. 2. Involuntary contraction Control by the autonomic nervous system The movement is undulating, it works slowly, since the nerve fiber does NOT come to each cell Slowly enter into action, but persist for a long time Does not have an accurate spatial orientation of cells 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiac The unit of organization is the muscle fiber - a multitude of myoblasts floating in the common cytoplasm (sarcoplasm). They share a common sarcolemma. Length about 40 -100 mm. Width from 7 mm. Voluntary contraction Controlled by the somatic nervous system Rapid contraction, rapid reaction, as each muscle fiber has a neuromuscular junction Quickly engaged but short-lived Clear orientation of muscle fibers
Striated Smooth 1. The unit of organization is the myocyte. The length is about 50 µm. Width from 6 µm. 2. Involuntary contraction Control by the autonomic nervous system The movement is undulating, it works slowly, since the nerve fiber does NOT come to each cell Slowly enter into action, but persist for a long time Does not have an accurate spatial orientation of cells 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Cardiac The unit of organization is the muscle fiber - a multitude of myoblasts floating in the common cytoplasm (sarcoplasm). They share a common sarcolemma. Length about 40 -100 mm. Width from 7 mm. Voluntary contraction Controlled by the somatic nervous system Rapid contraction, rapid reaction, as each muscle fiber has a neuromuscular junction Quickly engaged but short-lived Clear orientation of muscle fibers
 Between the muscle fibers are thin layers of loose fibrous connective tissue - endomysium. The collagen fibers of the outer sheet of the basement membrane are woven into it, which contributes to the unification of efforts during the contraction of myosymplasts. Thicker layers of loose connective tissue surround several muscle fibers, forming the perimysium and dividing the muscle into bundles. Several bundles are combined into more large groups separated by thicker connective tissue layers. The connective tissue surrounding the surface of the muscle is called the epimysium.
Between the muscle fibers are thin layers of loose fibrous connective tissue - endomysium. The collagen fibers of the outer sheet of the basement membrane are woven into it, which contributes to the unification of efforts during the contraction of myosymplasts. Thicker layers of loose connective tissue surround several muscle fibers, forming the perimysium and dividing the muscle into bundles. Several bundles are combined into more large groups separated by thicker connective tissue layers. The connective tissue surrounding the surface of the muscle is called the epimysium.
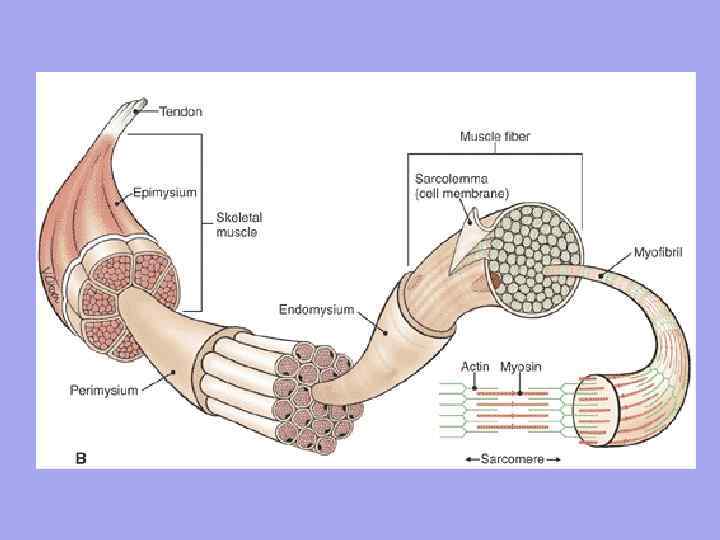
 In the muscle as an organ there is a connective tissue Endomysium - a thin connective tissue that surrounds each muscle fiber and small groups of fibers. Perimysium - covers larger complexes of muscle fibers and muscle bundles.
In the muscle as an organ there is a connective tissue Endomysium - a thin connective tissue that surrounds each muscle fiber and small groups of fibers. Perimysium - covers larger complexes of muscle fibers and muscle bundles.
 Significance of endomysium and perimysium 1. Vessels and nerves approach the muscle fiber through endomysium and perimysium. Form the stroma of the organ; 2. Muscle fibers are formed into bundles, bundles into muscle; 3. Since the endomysium is fused with the sarcolemma of the muscle fiber, therefore, the contracting muscle fiber can only stretch up to a certain limit
Significance of endomysium and perimysium 1. Vessels and nerves approach the muscle fiber through endomysium and perimysium. Form the stroma of the organ; 2. Muscle fibers are formed into bundles, bundles into muscle; 3. Since the endomysium is fused with the sarcolemma of the muscle fiber, therefore, the contracting muscle fiber can only stretch up to a certain limit
 Myofibrils in the fiber are surrounded by a shell - the sarcolemma, and immersed in a special environment - the sarcoplasm. Depending on the content of pigment and oxygen, the fibers are divided into white and red. White fibers are anaerobes, contain more myofibrils, less sarcoplasm. They start quickly, but they cannot work for a long time. Example: sternocleidomastoid, gastrocnemius muscles. Red fibers are thick fibers. There is a lot of myoglobin in the sarcoplasm and cytochrome in the mitochondria, but less myofibrils. Slow start, but work for a long time. Example: back muscles, diaphragm.
Myofibrils in the fiber are surrounded by a shell - the sarcolemma, and immersed in a special environment - the sarcoplasm. Depending on the content of pigment and oxygen, the fibers are divided into white and red. White fibers are anaerobes, contain more myofibrils, less sarcoplasm. They start quickly, but they cannot work for a long time. Example: sternocleidomastoid, gastrocnemius muscles. Red fibers are thick fibers. There is a lot of myoglobin in the sarcoplasm and cytochrome in the mitochondria, but less myofibrils. Slow start, but work for a long time. Example: back muscles, diaphragm.
 Each muscle has a network of blood vessels. Muscle contraction promotes blood flow. In a relaxed non-working muscle, most of the blood capillaries are closed to blood flow. When a muscle contracts, all blood capillaries immediately open.
Each muscle has a network of blood vessels. Muscle contraction promotes blood flow. In a relaxed non-working muscle, most of the blood capillaries are closed to blood flow. When a muscle contracts, all blood capillaries immediately open.

 Muscle structure Each muscle connects at one end to one bone (the beginning of the muscle), and at the other to the other (muscle attachment). In the muscle, they distinguish: head, abdomen, tail.
Muscle structure Each muscle connects at one end to one bone (the beginning of the muscle), and at the other to the other (muscle attachment). In the muscle, they distinguish: head, abdomen, tail.
 Motor nerve fibers approach each muscle fiber and sensory nerve fibers depart. The number of nerve endings in a muscle depends on the degree of functional activity of the muscles.
Motor nerve fibers approach each muscle fiber and sensory nerve fibers depart. The number of nerve endings in a muscle depends on the degree of functional activity of the muscles.
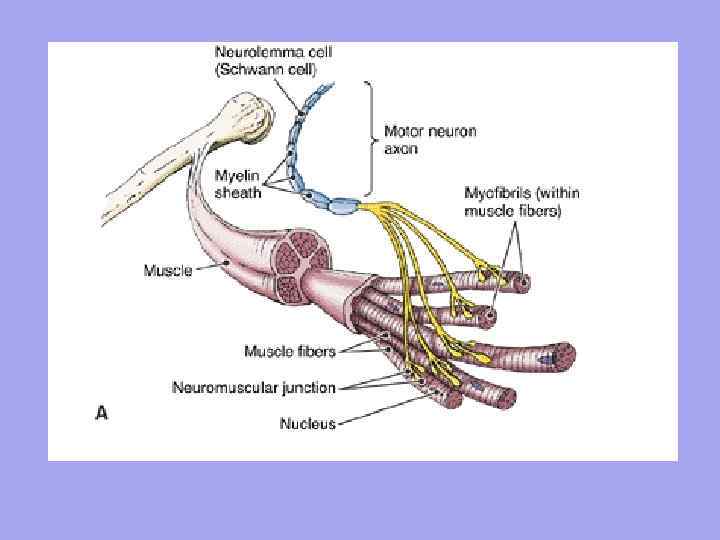
 Each muscle fiber is innervated independently and surrounded by a network of hemocapillaries, forming a complex called myon. A group of muscle fibers innervated by one motor neuron is called a motor unit. It is characteristic that the muscle fibers belonging to one motor unit do not lie side by side, but are located mosaically among the fibers belonging to other units.
Each muscle fiber is innervated independently and surrounded by a network of hemocapillaries, forming a complex called myon. A group of muscle fibers innervated by one motor neuron is called a motor unit. It is characteristic that the muscle fibers belonging to one motor unit do not lie side by side, but are located mosaically among the fibers belonging to other units.

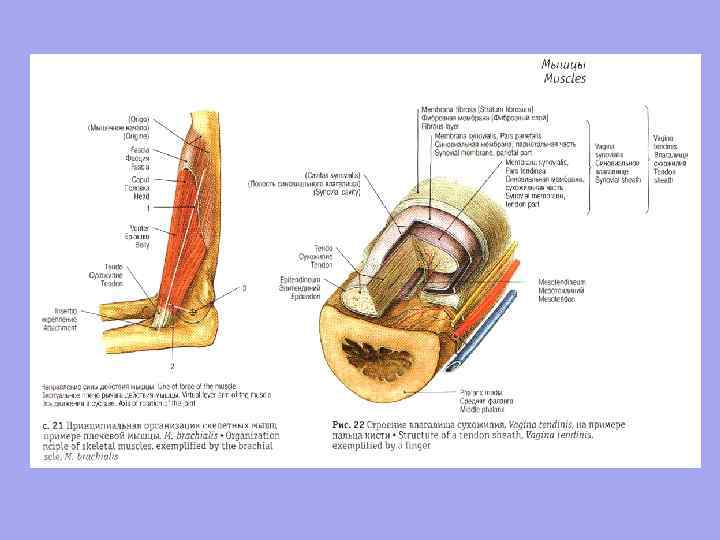
 A tendon is a dense fibrous connective tissue cord with which a muscle begins or attaches to the skeleton.
A tendon is a dense fibrous connective tissue cord with which a muscle begins or attaches to the skeleton.
 peritenonium collagen fibers of type IV endotenonium Collagen fibers of the tendon, intertwined with the collagen fibers of the periosteum, are woven into the main substance of the bone tissue, forming ridges, tubercles, tuberosities, depressions, depressions on the bones.
peritenonium collagen fibers of type IV endotenonium Collagen fibers of the tendon, intertwined with the collagen fibers of the periosteum, are woven into the main substance of the bone tissue, forming ridges, tubercles, tuberosities, depressions, depressions on the bones.
 Fascia are connective tissue collagen fibers with a small admixture of elastic fibers Superficial temporal fascia Deep thigh fascia
Fascia are connective tissue collagen fibers with a small admixture of elastic fibers Superficial temporal fascia Deep thigh fascia
 1. 2. 3. 4. 5. Fascia separate the muscles from the skin and eliminate the displacement of the skin during the movements of contracting muscles. Fascia conserve muscle contraction force by eliminating friction between muscles during contraction. Fascia stretch large veins under tension, as a result of which blood from the periphery is "sucked" into these veins. Fascia are important as barriers to the spread of infection and tumors. During operations, fasciae help determine the location of muscles, blood vessels, and viscera.
1. 2. 3. 4. 5. Fascia separate the muscles from the skin and eliminate the displacement of the skin during the movements of contracting muscles. Fascia conserve muscle contraction force by eliminating friction between muscles during contraction. Fascia stretch large veins under tension, as a result of which blood from the periphery is "sucked" into these veins. Fascia are important as barriers to the spread of infection and tumors. During operations, fasciae help determine the location of muscles, blood vessels, and viscera.
 Classification of muscles Skeletal muscles are diverse in shape, structure, position relative to the axes of the joints, etc., therefore they are classified in different ways.
Classification of muscles Skeletal muscles are diverse in shape, structure, position relative to the axes of the joints, etc., therefore they are classified in different ways.

 III. By functional features Static (strong) - a short abdomen and a long tendon. Muscles work with more force, but with a smaller range of motion. Dynamic (dexterous) - long muscle bundles, short tendons. Muscles work with less force but produce larger movements
III. By functional features Static (strong) - a short abdomen and a long tendon. Muscles work with more force, but with a smaller range of motion. Dynamic (dexterous) - long muscle bundles, short tendons. Muscles work with less force but produce larger movements
 Auxiliary apparatus of muscles Skeletal muscles have an auxiliary apparatus that facilitates their functioning. n n n Fascia; Bone-fascial sheaths; Synovial bags; Synovial tendon sheaths; muscle blocks; Sesamoid bones.
Auxiliary apparatus of muscles Skeletal muscles have an auxiliary apparatus that facilitates their functioning. n n n Fascia; Bone-fascial sheaths; Synovial bags; Synovial tendon sheaths; muscle blocks; Sesamoid bones.
 Anomalies in muscle development They are very common and are divided into three groups: 1. Absence of any muscle; 2. The presence of an additional muscle that does not exist in nature. 3. Additional bundles of existing muscle.
Anomalies in muscle development They are very common and are divided into three groups: 1. Absence of any muscle; 2. The presence of an additional muscle that does not exist in nature. 3. Additional bundles of existing muscle.
 Malformations Underdevelopment of the sternocleidomastoid muscle - Torticollis Underdevelopment of the diaphragm. Cause of diaphragmatic hernia. Underdevelopment of the deltoid and trapezius muscles - Deformity of the shoulder girdle and shoulder
Malformations Underdevelopment of the sternocleidomastoid muscle - Torticollis Underdevelopment of the diaphragm. Cause of diaphragmatic hernia. Underdevelopment of the deltoid and trapezius muscles - Deformity of the shoulder girdle and shoulder


 I. Shape: Fusiform; ribbon-like; Flat wide; jagged; Long; n n n Square; triangular; round; deltoid; soleus, etc.
I. Shape: Fusiform; ribbon-like; Flat wide; jagged; Long; n n n Square; triangular; round; deltoid; soleus, etc.



 II. In the direction of muscle fibers With straight parallel fibers; With transverse; With circular; Pinnate: A. Unipinnate; bipinnate; C. Multipinnate. b.
II. In the direction of muscle fibers With straight parallel fibers; With transverse; With circular; Pinnate: A. Unipinnate; bipinnate; C. Multipinnate. b.

 IV. By function: Leading; outlet; flexors; extensor; Pronators; n n Arch supports; Straining; Muscles are synergists; Muscles are antagonists.
IV. By function: Leading; outlet; flexors; extensor; Pronators; n n Arch supports; Straining; Muscles are synergists; Muscles are antagonists.
 V. In relation to the joint: Single-joint; biarticular; Polyarticular.
V. In relation to the joint: Single-joint; biarticular; Polyarticular.
Non-isolated muscular system
Single skin-muscle bag
The appearance of striated muscle tissue
Separation of muscle strands into myotomes
Development of muscle groups
Limb muscle development (habitat change)
Diaphragm Development
Development of all muscle groups - performance of differentiated movements
2 Ontogeny of the muscular system: sources and timing of development.
Myotome derivatives: back muscles develop from the dorsal region
from the ventral - the muscles of the chest and abdomen
Mesenchyme - limb muscles
I visceral arch (VD) - masticatory muscles
II VD - mimic muscles
III and IV VD - muscles of the soft palate, pharynx, larynx, upper esophagus
V VD - sternocleidomastoid and trapezius muscles
From the occipital myotomes - the muscles of the tongue
From the anterior myotomes - the muscles of the eyeball
Muscles develop from mesoderm. On the trunk, they arise from the primary segmented mesoderm - somites: 3-5 occipital, 8 cervical, 12 thoracic, 5 lumbar, 5 sacral, 4-5 coccygeal.
Each somite is subdivided into sclerotome, dermatome and myotome- from it the muscles of the body develop. Somites appear early, when the length of the embryo is 10-15 mm.
From dorsal parts of myotomes arise deep, own(autochthonous) muscles of the back, from ventral- deep muscles of the chest and abdomen. They are laid, develop and remain within the body - therefore they are called autochthonous (local, native). Very early, myotomes bind to nervous system and each muscle segment corresponds to a nerve segment. Each nerve follows the developing muscle, grows into it, and, until it is differentiated, subordinates to its influence.
In the process of development, part of the skeletal muscles moves from the trunk and neck to the limbs - trunk-fugal muscles: trapezius, sternocleidomastoid, rhomboid, levator scapula, etc. Part of the muscles, on the contrary, is directed from the limbs to the trunk - truncal muscles: latissimus dorsi, pectoralis major and minor, psoas major.
Muscles of the head facial and chewing, supra- and hyoid muscles of the neck develop from non-segmented ventral mesoderm, which is part of the visceral (gill) arches. They are called visceral and, for example, chewing muscles develop on the basis of the first visceral arc, and mimic - the second. However, the muscles of the eyeball and tongue develop from the occipital myotomes of the segmented mesoderm. The deep anterior and posterior muscles of the neck also arise from the occipital cervical myotomes, while the superficial and middle group muscles in the anterior region of the neck develop on the basis of the non-segmented mesoderm of the visceral arches.
3 Muscle: definition, structure.
Muscle(muscle) - an organ built from muscle fibers (cells), each of them has a connective tissue sheath - endomysium. Another fibrous sheath unites muscle fibers into bundles - perimysium, and the entire muscle is enclosed in a common fibrous sheath formed by fascia - epimysium. Between the bundles are vessels and nerves that supply the muscle fibers.
At the macro level, skeletal muscle has:
· abdomen(venter) - the fleshy part of the body, which occupies its middle;
· tendon(tendo) related to the distal end, it can be in the form of an aponeurosis, tendon bridges, long bundles of longitudinal fibrous fibers;
· head, constituting the proximal part;
tendon and head are attached at opposite ends of the bones.
The proximal tendon or head of the muscle - the beginning of the muscle on the bone is closer to the median axis of the body - this is a fixed point (punctum fixum) (usually coincides with the beginning of the muscle). The distal tendon, "tail" - the end of the muscle lies distally on the bone and, being the point of attachment, is called a mobile point (punctom mobile). When the muscle contracts, the points approach each other, and when the position of the body changes, they can change places.
The tendons are different in shape: thin long tendons have limb muscles; the muscles involved in the formation of the walls of the abdominal cavity have a wide flat tendon located between the two bellies - a tendon stretch or aponeurosis.
4 Classification of muscles by origin, structure, form and function.
A. Vesalius, an anatomist of the Renaissance, designated the muscles with numbers, but now they are classified according to other principles.
Origin:
- from dorsal parts of myotomes arise deep, own(autochthonous) back muscles
- from ventral- deep muscles of the chest and abdomen, they are laid, develop and remain within the body - therefore they are called autochthonous (local, native).
By function, they are distinguished:
muscles- antagonists, such as: flexors and extensors, adductors and abductors, supinators and pronators - such muscles act in opposite directions;
muscles- synergists- act in the same direction, reinforcing each other; with a number of movements, antagonist muscles also act as synergists, for example, when performing circular movements;
major and accessory muscles.
By location:
external and internal
superficial and deep
medial and lateral
Form and structure:
Fusiform muscles (musculi fusiformes) - yavl. long levers (biceps brachii)
broad muscles - participate in the formation of the walls of the body (rectus abdominis)
One-, two- and multi-pinnate muscles - depending on whether muscle bundles lie on one side of the tendon or on two or more sides, for example, the multi-pinnate deltoid muscle.
Muscles whose shape corresponds to a certain geometric figure, for example, rhomboid large and small, trapezoid, square, circular, straight, thin;
Muscles with several heads or bellies: two-, three-, quadriceps muscles of the limbs; digastric on the neck;
Muscles whose names reflect the direction of the fibers: transverse, longitudinal, oblique;
Muscles whose names reflect the function: extensor, flexor, adductor, abductor, lifting, lowering, compressing, etc.;
Muscles that are large in area and length: wide and latissimus, large and small, long and short;
One-, two- and multi-joint muscles, depending on how many joints the muscles act on, there are muscles that do not act on the joint at all.
They also distinguish:
- smooth muscles (involuntary muscles) - develop from the visceral sheet of the splashnotome, are located in the wall internal organs, contract spontaneously, innervated by the autonomic nervous system;
- striated muscles - skeletal (arbitrary muscle, developed from myotomes - arr. skeletal musculature, inerv. - somatic nervous system) and cardiac (involuntary musculature, has a cross-striped structure, but consists of departments. cells - cardiomyocytes, nerve - vegetative nervous system)
Skeletal muscles perform overcoming, yielding work, which provides muscular dynamics of the body, holding- myostatic work.
Somatic and visceral muscular system, its phylo-ontogenesis. Subcutaneous muscles. Skeletal muscles. The structure of a muscle as an organ. Muscle classification. Muscle accessories.
myology(Myologia) is a branch of the anatomy of domestic animals that studies the structure of the muscular system. Muscle tissue, which forms the basis of this system, carries out all motor processes in the animal body. Thanks to it, the body is fixed in a certain position and moves in space, respiratory movements of the chest and diaphragm, eye movement, swallowing, motor functions of internal organs, including the work of the heart, are carried out.
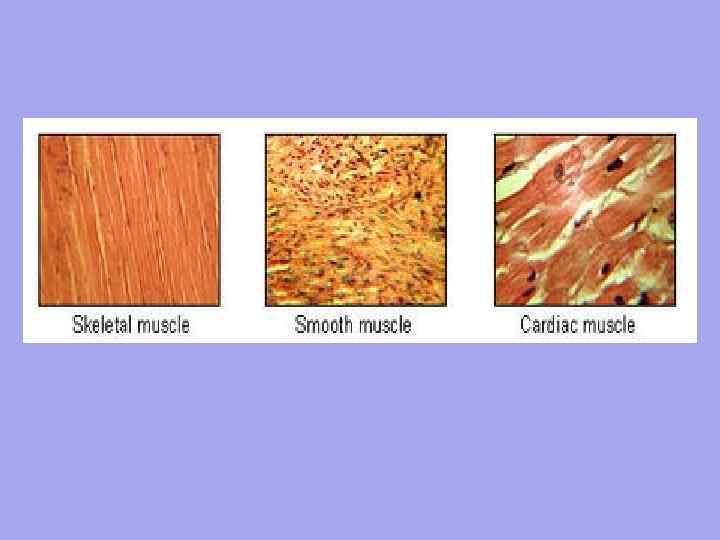
Muscle has special contractile organelles - myofibrils . myofibrils, consisting of thin protein filaments (myofilaments), may be unstriated or striated (striated). Accordingly, unstriated and striated muscle tissue is distinguished.
1) Non-striated muscle tissue consists of cells (smooth myocytes) of a fusiform shape. These cells form muscle layers in the walls of blood and lymph vessels, in the walls of internal organs (stomach, intestines, urinary tract, uterus, etc.). The length of the cells ranges from 20 µm (in the wall of a blood vessel) to 500 µm (in the wall of the uterus of a pregnant cow), the diameter is from 2 to 20 µm. In functional terms, unstriated muscle tissue has a number of features: it has great strength (for example, significant masses of food constantly move in the intestines), has low fatigue, slow contraction and rhythm of movements (in the intestinal wall, unstriated muscle tissue contracts 12 times per minute, and in spleen - only 1 time).
2) Striated muscle tissue is characterized by the presence of striated myofibrils, has 2 varieties.
A) striated cardiac muscle tissue consists of elongated cells (cardiomyocytes) is square. Their ends, connecting with each other in chains, form the so-called functional muscle "fibers" with a thickness of 10-20 microns. Closely communicating with each other, functional muscle "fibers" form the muscular membrane of the heart ( myocardium), the constant and rhythmic contractions of which set the blood in motion.
B) Striated skeletal muscle tissue, unlike cardiac tissue, does not consist of cells, but of multinuclear muscle formations (myosymplasts) of a cylindrical shape. The length of myosymplasts varies from a few millimeters to 13-15 cm, the diameter is from 10 to 150 microns. The number of cores in them can reach several tens of thousands. Myosymplasts (also called "muscle fibers") form skeletal muscles and are part of some organs (tongue, pharynx, larynx, esophagus, etc.). In functional terms, skeletal muscle tissue is easily excitable and contracts faster than non-striated muscle (for example, under normal conditions, skeletal muscle contracts within 0.1 s, and non-striated muscle within a few seconds). But, unlike the smooth (non-striated) muscles of the internal organs, skeletal muscles tire faster.
Muscular system depending on the structural features, the nature of the motor function and innervation, they are divided into somatic and visceral.
somatic muscular system makes up 40% of body weight and is built from myosymplasts. It is voluntary and innervated by the somatic nervous system. Somatic muscles contract quickly, vigorously, but they tire for a short time and quickly. This type of reduction is called tetanic and it is characteristic of somatic musculature. It includes:
1) subcutaneous muscles that have no connection with the skeleton and are attached to the skin; their contractions cause twitching of the skin and allow it to gather into small folds;
2) skeletal muscles, which are fixed on the skeleton;
3) diaphragm - a dome-shaped muscle that separates the chest cavity from the abdominal cavity;
4) muscles of the tongue, pharynx, larynx, auricle, eyeball, middle ear, esophagus and external reproductive organs.
visceral muscular system makes up 8% of body weight and is built from smooth myocytes. It is involuntary and is innervated by the autonomic nervous system. Smooth muscles contract slowly, for a long time and do not require a lot of energy. This type of reduction is called tonic and it is characteristic of the visceral musculature, which forms muscle bundles, layers and membranes of internal organs.
Phylo-ontogeny of the muscular system
In the phylogeny of chordates, the muscular system successively passes through a series of stages.
At the lancelet it is represented by a paired longitudinal muscle (right and left), which runs along the body and is divided by connective tissue septa (myoseptae) into short straight muscle bundles (myomeres). This (segmental) division of a single muscle layer is called metamerism.
With an increase in mobility, isolation of the head and development of the limb (in the form of fins) in fish there is a division of the longitudinal muscle by a horizontal septum into dorsal and ventral muscles, as well as
Isolation of the muscles of the head, trunk, tail and fins.
With access to land and an increase in the variety of movements in amphibians and reptiles there is a division of the dorsal muscle, as well as the ventral one, into two cords: lateral (transverse costal muscle) and medial (transverse spinous m.). In addition, in reptiles, subcutaneous muscles appear for the first time from the lateral cord, which are attached to the skin.
In more highly organized animals ( birds and mammals) further differentiation of the muscular system occurs: the lateral and medial strands, each of them, are divided into two layers (superficial and deep). In addition, the diaphragm appears for the first time in mammals.
Phylogeny of the muscular system.
| chordates | Muscular system | |||||||
| Lancelet | longitudinal muscle | |||||||
| Fish | Dorsal | Ventral | ||||||
| Amphibians, reptiles | Lateral | Medial | Lateral | Medial | ||||
| Birds, mammals | Power | Depth | P | G | P | G | P | G |
In ontogenesis, the muscular system mainly develops from the myotomes of the mesoderm, with the exception of some muscles of the head and neck, which are formed from the mesenchyme (trapezoid, brachiocephalic).
At the beginning, a muscular longitudinal cord is formed, which immediately differentiates into dorsal and ventral layers; further, each of them is divided into lateral and medial layers, which, in turn, differentiate into superficial and deep layers, the latter give rise to certain muscle groups. For example, the iliocostal muscles develop from the surface layer of the lateral layer, and the longest muscles of the back, neck, and head develop from the deep layer of the lateral layer.
Subcutaneous muscles - musculi cutanei
Subcutaneous muscles are attached to the skin, fascia and have no connection with the skeleton. Their contractions cause the skin to twitch and allow it to gather into small folds. These muscles include:
1) Subcutaneous muscle of the neck - m. Cutaneus colli (especially strongly developed in dogs). It goes along the neck, closer to its ventral surface and passes to the front surface to the muscles of the mouth and lower lip.
2) Subcutaneous muscle of the scapula and shoulder (scapulohumeral) - m. Cutaneus omobrachialis. It covers the area of the scapula and partially the shoulder. Well expressed in horses and cattle.
3) Subcutaneous muscle of the body - m. Cutaneus trunci. It is located on the sides of the chest and abdominal walls and caudally gives the bundles to the knee crease.
4) In females, in the area of \u200b\u200bthe mammary glands, there are cranial and caudal muscles of the mammary gland (mm. Supramammilaris cranialis et caudalis), which give folding to the skin and help to remove milk. Strongly developed in carnivores.
Males in this area have cranial and caudal preputial muscles (mm.preputialis cranialis et caudalis), which provide folding of the prepuce and act as its sphincter.
skeletal muscles
Skeletal muscles are the active part of the musculoskeletal system. It consists of skeletal muscles and their accessories, which include fascia, synovial bags, synovial tendon sheaths, blocks, sesame bones.
There are about 500 skeletal muscles in the body of an animal. Most of them are paired and are located symmetrically on both sides of the animal's body. Their total weight is 38-42% of body weight in a horse, 42-47% in cattle, and 30-35% in pigs.
The muscles in the animal's body are not arranged randomly, but naturally, depending on the action of the animal's gravity and the work performed. They exert their effect on those parts of the skeleton that are movably connected, i.e. muscles act on joints, syndesmoses.
The main places of attachment of muscles are bones, but sometimes they are attached to cartilage, ligaments, fascia, skin. They cover the skeleton so that the bones only in some places lie directly under the skin. Fastening on the skeleton, as on a system of levers, the muscles cause various movements of the body during their contraction, fix the skeleton in a certain position and give shape to the body of the animal.
The main functions of skeletal muscles:
1) The main function of the muscles is dynamic. When contracting, the muscle shortens by 20-50% of its length and thereby changes the position of the bones associated with it. Work is done, the result of which is movement.
2) Another function of the muscles is static. It manifests itself in fixing the body in a certain position, in maintaining the shape of the body and its parts. One of the manifestations of this function is the ability to sleep standing up (horse).
3) Participation in the metabolism and energy. Skeletal muscles are "sources of heat", since during their contraction about 70% of the energy is converted into heat and only 30% of the energy provides movement. About 70% of the body's water is retained in skeletal muscles, which is why they are also called "sources of water." In addition, adipose tissue can accumulate between the muscle bundles and inside them (especially when fattening pigs).
4) At the same time, during their work, skeletal muscles help the work of the heart, pushing venous blood through the vessels. In experiments, it was possible to find out that skeletal muscles act like a pump, ensuring the movement of blood through the venous bed. Therefore, skeletal muscles are also called "peripheral muscle hearts."
The structure of the muscle from the point of view of a biochemist
Skeletal muscle is made up of organic and inorganic compounds. Inorganic compounds include water and mineral salts (salts of calcium, phosphorus, magnesium). Organic matter is mainly represented by proteins, carbohydrates (glycogen), lipids (phosphatides, cholesterol).
Table 2. Chemical composition skeletal muscle
The chemical composition of skeletal muscles is subject to significant age and, to a lesser extent, species, breed and sex differences, which is primarily due to the unequal water content in them (with age, the percentage of water decreases).


