ті, що розташовуються на дистальному кінці протиспинкової поверхні кісток п'ясті, плюсни та дистальних фаланг пальців (див. скелет). До сесамоподібних кісток відносяться колінна чашка та додаткова кістка зап'ястя.
КОРОТКІ ВІДОМОСТІ ПО ФІЛО ТА ОНТОГЕНЕЗ МУСКУЛАТУРИ
Філогенетичні перетворення. М'язові елементи в ряду раз-
вития живих істот з'являються рано у кишковопорожнинних. Вони ще не відокремлені самостійні морфологічні одиниці, а є лише скоротливими м'язовими елементами епітеліальних клітин. Надалі вони відокремлюються від епітелію, утворюючи кілька шарів гладком'язових клітин, тісно з'єднаних зі шкірою, у результаті утворюється так званий шкірно-м'язовий мішок (плоскі черв'яки). Джерелом утворення м'язових клітин стає мезодерма.
З появою вторинної порожнини тіла відбувається поділ мускулатури на соматическую, що входить до складушкірно-м'язового мішка, і вісцеральну, що оточує кишечник та судини. Незважаючи на такий поділ, вона може бути або вся гладка (кільчасті черв'яки), або вся поперечносмугаста (комахи). Це свідчить про те, що у філогенезі поперечнополосата мускулатура майже не відрізняються від гладкої ні за походженням, ні за функцією. З подальшим ускладненням організації соматична і вісцеральна мускулатура розвиваються дивергентно, дедалі більше розходячись один від одного структурно та функціонально.
У примітивних хордових (ланцетник, круглороті) вся соматична мускулатура розвивається із сомітів мезодерми і є поперечнополосатою. Вона являє собою парну праву і ліву поздовжні м'язи, що йдуть вздовж усього тіла, поділені сполучнотканинними перегородками.Міосептами на ряд міомерів - коротких сегментів із прямих м'язових пучків. Такий (сегментарний) поділ єдиного м'язового пласта називається метамерією (рис. 73).
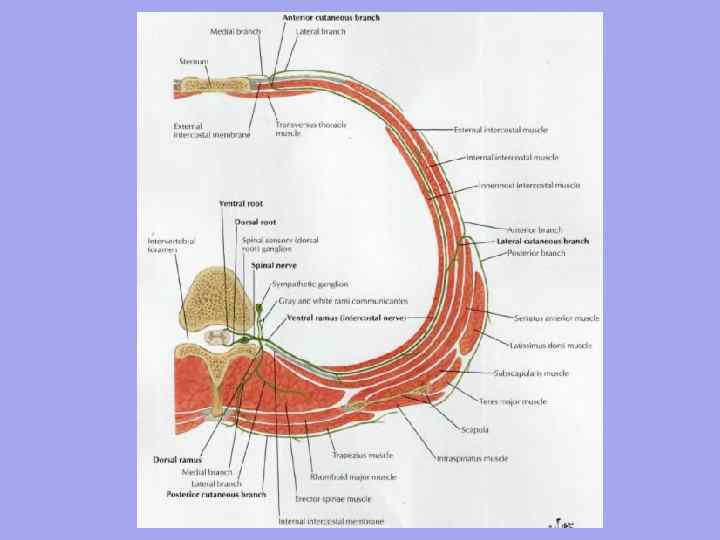
З відокремленням голови та розвитком кінцівок (у вигляді плавників) диференціюється і мускулатура. Поздовжній м'яз у риб ділиться горизонтальною септою надорсальну та вентральні м'язи.Вони іннервуються відповідно дорсальними та вентральними гілками спинномозкових нервів. Іннервація ця зберігається за всіх подальших перетвореннях м'язів. Дорсальні та вентральні поздовжні м'язи у зв'язку з одноманітністю рухів первинних тварин мають міомерну будову. Кожному міомеру зазвичай відповідає свій хребець і парний спинномозковий нерв. У вищих риб (оселедеві та інших.) можна побачити їх поздовжнє розщеплення деякі шари. Відокремлюється і мускулатура плавників, однак у порівнянні з м'язами стовбура тіла вона розвинена слабо, оскільки основне навантаження під час руху у водних тварин лягає на хвіст і тулуб.
Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
Мал. 73. М'язи тіла хордових тварин:
А – ланцетник; 5-риба; В – хвостата амфібія; Г-рептилія; 1 – міомери (міотоми); 2-міосепти; 3- дорсальна м.. тулуба; 4- поздовжня бічна перегородка; 5 – дорсальна м. хвоста; 6 - поверхневий стискувач; 7- трапецієподібна м.; 8 – вентральна м. тулуба; 9 – вентральна м. хвоста; 10 – мм. грудної кінцівки; 11 - найширша м, спини; 12, 13, 14 – вентральні мм. (12 - коса зовнішня, 13-коса внутрішня, 14 - пряма); 15 – мм. тазової кінцівки.
З виходом на сушу та збільшенням різноманітності рухів зростає розчленування м'язових пластів на окремі м'язи як вздовж, так і впоперек. У цьому метамерність поступово зникає. Вона добре простежується в м'язах риб, помітна ще й у амфібій, слабо - у рептилій. У ссавців збережена лише у глибоких шарах, де короткі м'язи з'єднують між собою елементи двох сусідніх кісткових сегментів (міжостисті, міжпоперечні, міжреберні м'язи).
Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
Насамперед метамерія починає зникати в черевному відділі тулуба, де вже в амфібій окремі міомери зливаються з утворенням широких пластинчастої форми черевних м'язів. Разом з цим йде поздовжнє розщеплення м'язової черевної стінки із формуванням чотиришарового черевного преса. У дорсальній мускулатурі тулуба амфібій можна вичленувати два тяжи: латеральний і медіальний, метамерія яких загасається лише в шийному відділі, де відокремлюються самостійні м'язи
У рептилій м'язові пучки латерального та медіального м'язових тяжів набувають різної спрямованості. Міомерія зберігається лише у глибоких шарах. Що ближче до голови, то ясніше роздроблення дорсальних тяжів на окремі м'язи.
У ссавців соматична мускулатура диференційована найбільше. У дорсальній мускулатурі формується 4 шари за рахунок розшарування латерального та медіального м'язових тяжів. При цьому спостерігається чітка закономірність: чим глибше знаходиться м'яз, тим краще виражена його метамерія; що ближче до зовнішньої поверхні тіла лежить м'яз, то більше вона втрачає метамерію, поширюючись по тілу широким пластом. Розчленованість дорсальної мускулатури зростає і в краніальному напрямку, що пов'язано зі ступенем рухливості хребта. Якщо в області крижів - самого нерухомого відділу стовбурового скелета
- Дорсальні м'язи абсолютно не розчленовані, то в ділянці холки, і особливо шиї, м'язові комплекси складаються з великої кількості самостійних м'язів.
Вентральна мускулатура стовбурової частини тіла також має 4 шари, щоправда, не скрізь виражені повною мірою. У грудній клітці це внутрішні та зовнішні міжреберні, прямі та поперечні грудні м'язи, у попереково-черевному відділі - м'язи черевного преса.
Локомоторна функція мускулатури хвоста з виходом на сушу стає все менше і у ссавців повністю втрачається. Це призводить до значного зменшення маси м'язів за збереження високого ступеня їх диференційованості у зв'язку з рухливістю хвоста.
Кінцівки наземних хребетних походять з кистеперого плавця, дуже рухливого, з добре розвиненим скелетом та сильною мускулатурою (латимерія). Метамерність мускулатури кінцівок, добре помітна у променеперих риб, у філогенезі втрачається дуже рано, особливо з виходом на сушу. З перетворенням кінцівки на складний важіль, що підтримує і пересуває тіло тварини по суші, відбувається відокремлення великої кількості м'язів.
Для примітивних чотирилапих характерне відходження плечової та стегнової кісток убік та вгору від пояса. За такого розташування відділів кінцівок потрібні великі витрати м'язової енергії підтримки тіла на вису. На грудній кінцівці найбільше навантаження падає на коракоїдну кістку, до якої в результаті кріпиться основна маса мускулатури плечового та ліктьового суглобів.
Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
Пристосування до швидкого бігу, маніпуляції грудною кінцівкою і здатність відпочивати стоячи, що розвинулися у ссавців, супроводжувалися поворотом кінцівки із сегментальної до сагітальної площини розкриттям суглобів і все більш високим підніманням тіла над землею. При цьому змінювалися умови дії сили тяжіння та роботи м'язів при стоянні та русі тварини. У копитних пристосування кінцівок до швидкого поступального руху і економного витрачання м'язової енергії при стоянні призвело до втрати різноманітності руху. Виявилося це ще більшої редукції плечового пояса (зникнення ключиці) і випрямлення вільної кінцівки. Плечовий пояс втратив кістковий зв'язок з осьовою частиною тіла і набув великої площі опори за допомогою м'язів, що зв'язали його з головою, шиєю, холкою, спиною та грудною клітиною. Так що мускулатура кінцівок по масі почала переважати над мускулатурою тулуба. М'язи поясів та проксимальних відділів кінцівок значною мірою покривають зверху тулубну мускулатуру та частково її витісняють. Розвиток м'язів дистальних ланок багато в чому визначається особливостями механіки руху та екології тварини (ходіння, повзання, стрибання, риття тощо). У копитних у зв'язку з редукцією пальців, випрямленням суглобів сталося зменшення кількості до ускладнення будови м'язів дистальних ланок кінцівок.
І нарешті, найбільш поверхневим і найменш розчленованим м'язовим пластом є підшкірна мускулатура - частина соматичної мускулатури, що вперше виникла у рептилій. У ссавців вона сильно розвинена, особливо у тварин, які можуть згортатися (їжак, броненосець). З домашніх тварин вона добре розвинена у коня і має вигляд широких пластів, що лежать під шкірою в області шиї, холки, лопатки, грудей та черева (див. рис. 72). На голові підшкірна мускулатура входить у тісний зв'язок із вісцеральною та входить складовоюу м'язи обличчя, повік, носа, вушної раковини.
Складні перетворення на м'язах голови відбуваються паралельно зі складними филогенетическими перетвореннями черепа. В результаті соматична мускулатура в ділянці голови значною мірою витісняється вісцеральною мускулатурою, що оточує головну кишку. Соматична мускулатура голови вже у риб, представлена лише м'язами ока та деякими над-і поджаберними м'язами з поздовжнім напрямом м'язових волокон (беруть участь у дихальних рухах зябрового апарату).
Вісцеральна мускулатура, що оточує головний кінець кишкової трубки, зазнала значного диференціювання, набула властивостей поперечносмугастої м'язової тканини, але зберегла свій кільцевий напрямок волокон. Нею утворені кільцеві м'язові пласти щелепної, під'язикової та зябрових дуг, на базі яких розвивається основна маса м'язів голови: щелепні, під'язичні, зяброві, деякі м'язи плечового пояса з хапальною, жувальною та іншими функціями.

Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
У ссавців соматична мускулатура голови представлена м'язами ока, середнього вуха, язика та деякими м'язами під'язикової кістки. Вісцеральна мускулатура утворює мімічні (лицьові) та жувальні (щелепні) м'язи.
І нарешті, тільки у ссавців є м'язова грудобрюшна перешкода - діафрагма.
Онтогенетичний розвиток. Соматична мускулатура переважно походить з міотомів сомітів мезодерми (рис. 74). В області голови з трьох міотомів утворюються м'язи очного яблука. Передні завушні міотоми зникають, та якщо з задніх (потиличних) розвивається під'язична мускулатура. Вісцеральна мускулатура голови мезенхімного походження. Шийні, грудні, поперекові, крижові та хвостові міотоми закладаються відповідно до кількості метамерних сегментів тіла. Вони розростаються в дорсальному та вентральному напрямках і дають початок всій соматичній мускулатурі шиї, тулуба та хвоста. М'язи кінцівок утворюються шляхом виростів вентральних ділянок міотомів, до яких приєднується клітинний матеріал, що виселяється з парієтального листка спланхнотома мезодерми. Закладка мускулатури дещо відстає від закладення скелета і певною мірою залежить від нього.
Мал. 74. Метамерна закладка мускулатури у зародка ссавця міотоми:
1- потиличні. 2 – шийні, 3 – грудні. 4 - поперекові, 5 - крижові, 6 - хвостові.
У зародковий період, з 20-22-го дня розвитку, у великого рогатої худобиу міотомах відбувається розмноження міобластів. У передплідний період починається анатомічна диференціювання: відокремлюються м'язи та групи м'язів. Паралельно з цим, але значно довше йде гістогенез м'язової тканини. Міобласти зливаються в м'язові трубочки, у них з'являються міофібрили. Анатомічна диференціювання в основному закінчується в передплідному періоді - до 50-55-го дня. Закладка та диференціювання м'язів йде у певній послідовності. Раніше за інших закладається осьова мускулатура. У ньому диференціювання йде від головного кінця до хвостового. При цьому глибокі м'язи диференціюються раніше
Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
поверхневих. У процесі диференціювання м'язів у них вростають відповідні черепноабо спинномозкові нерви. Цей зв'язок встановлюється дуже рано і зберігається протягом усього життя. Закладки кінцівок з'являються у вигляді валикоподібних потовщень біля вентральних ділянок від 5-го шийного до 1-го грудного міотома - зачаток грудної кінцівки та від 1-го поперекового до 3-го крижового міотома - зачаток тазової кінцівки. Незабаром валики стягуються і набувають вигляду сплющених конічних виростів - нирок. Утворення м'язів на грудній кінцівці у зародка теля починається з 32-го дня, а на задній - з 34-го дня ембріонального розвитку. Раніше за інших закладаються м'язи поясів, потім вільної кінцівки, де процес поширюється від проксимальних ланок до дистальних. Як і осьової частини тіла, диференціювання глибоких м'язів йде раніше, поверхневих - пізніше. На латеральному боці кінцівки закладаються екстензори, абдуктори та супінатори, на медіальній – флексори, аддуктори та пронатори. Черевці м'язів закладаються раніше за сухожилля. До кінця передплідного періоду м'язи кінцівок виявляються анатомічно сформовані, а гістологічно незрілі - складаються з м'язових трубочок, що лежать пучками. Протягом плодового періоду продовжується гістологічна диференціювання м'язів: збільшуються число та розміри м'язових трубочок, відбувається перетворення трубочок на м'язові волокна, наростає в них кількість міофібрил; оформляється ендомізій та перимизій м'язів, розвиваються капілярні мережі, оформляються пучки I, II та III порядків.
В результаті анатомо-гістологічного диференціювання з дорсальних ділянок міотомів утворюється дорсальна мускулатура хребетного стовпа, що лежить над тілами хребців. Вона іннервується дорсальними гілками спинномозкових нервів. З вентральних ділянок міотомів утворюється вентральна мускулатура хребетного стовпа, що лежить під тілами хребців, мускулатура. грудної клітки, черевної стінки та діафрагми. З м'язових бруньок розвивається вся мускулатура кінцівок.
У У процесі органогенезу відбувається відокремлення м'язів за довжиною, товщиною, дробленням або злиттям, утворенням комплексних і багатороздільних м'язів, формуванням їх перистої структури. У ранній плодовий період швидше ростуть м'язи тулуба, а в пізній - м'язи кінцівок, особливо їх дистальних ланок - лапи.
У копитних тварин до народження повністю сформований апарат руху, який відразу ж починає функціонувати: через кілька годин новонароджене теля, ягня, лоша, порося може слідувати за матір'ю. Однак це не означає, що в локомоторному апараті завершено процеси зростання та диференціювання. Вони продовжуються до віку морфофізіологічної зрілості, а адаптивна перебудова апарату руху відбувається протягом усього життя.
Постнатальне зростання мускулатури. Після народження продовжується інтенсивне зростання мускулатури, яка за швидкістю зростання обганяє скелет. Особливо посилено цей процес протікає в перші два місяці після народження.
Вракін В.Ф, Сидорова М.В. |
МОРФОЛОГІЯ СІЛЬСЬКОГОСПОДАРСЬКИХ ТВАРИН |
ня. Наступні піки зростання у великої рогатої худоби припадають на 6-й та 12-й місяці життя, у овець – на 3-й та 9-й місяці. Осьова мускулатура росте швидше, ніж м'язи кінцівок, особливо з настанням статевої зрілості. У новонароджених телят маса осьової мускулатури становить 46%. а у 14-місячних – 53%. На кінцівках велика швидкість зростання м'язів у проксимальних ланках (проти дистальними). На грудній кінцівці вони ростуть дещо інтенсивніше, але швидше завершують зростання, ніж м'язи тазової кінцівки. Розгиначі ростуть швидше за згиначів, причому періоди підйому швидкості їх зростання не збігаються.
З віком у м'язах зменшується кількість м'язових волокон на одиницю площі та в первинних м'язових пучках, так як поряд з потовщенням м'язових волокон (приблизно в 15-20 разів) відбувається проростання м'язів сполучною тканиною, вона стає щільнішою, м'язові пучки. I порядку включають менше волокон. Тим не менш відносна кількість сполучної тканини в м'язах з віком знижується, а м'язової
Збільшується. Так, за 18 місяців у бичків кількість сполучної тканини збільшується у 8 разів, а м'язової – у 17 разів. Змінюється і хімічний склад: збільшується кількість протеїну та жиру, менше стає води. До кожного типу м'язів характерна своя динаміка хімічних показників.
Не тільки групи м'язів, а й кожен м'яз має власний характер зростання, що пов'язано як з особливостями її внутрішньої структури так і функціонування. Найвищі темпи зростання м'язів динамічного типу. Нерівномірність зростання м'язів багато в чому визначає зміна пропорцій та статей тіла.
Вплив внутрішніх та зовнішніх факторів на зростання м'язів. Спосіб життя тварини, спосіб видобутку та характер корму накладають відбиток на зростання та диференціювання мускулатури. Так, у свиней більше розвивається дорсальна мускулатура, особливо шиї. У коня краще, ніж рогатої худоби, розвинені жувальні м'язи. Черевні м'язи, навпаки, більш розвинені у худоби.
На характер зростання м'язів накладає відбиток та стать тварини. При однаковій вгодованості у бичків м'язи розвинені краще і становлять у туші більший відсоток, ніж у телиць та бичків-кастратів. До того ж зростання мускулатури у бичків триває довше, отже від них зрештою можна отримати більше м'яса. У бичків більш розвиненою виявляється мускулатура шиї, холки та плечового пояса (що важливо для сили тварини при встановленні ієрархії у стаді). У телиць розвиненіша мускулатура черевного преса і задньої половини тіла. Кастрати за характером зростання мускулатури наближаються до телиць, а за зростанням найдовших і напівостистих м'язів відстають від тварин обох статей. У бичків у м'язах менше жирових включень, а у телиць і кастратів тонші за м'язові волокна, добре виражена мармуровість м'яса.
Спостерігаються деякі відмінності в темпах зростання та розвитку мускулатури між породами з різним напрямом продуктивності. Для скоростиглих порід характерна висока енергія росту, зате у пізньостиглих
Складне філогенетичний розвиток м'язів тулуба, шиї та голови обумовлено тим, що вони походять з двох зачатків: одна частина є власною мускулатурою тулуба, інша – закладається у бічних пластинках, що належать до кишкової мускулатури, пов'язаної з мезенхімою зябрового апарату.
Для правильного уявлення еволюції м'язів розберемо їх зачатки.
Філогенез м'язів тулуба. Нижчі хребетні, подібно до ланцетника, мають парні м'язи, розташовані з боків тулуба. Бічні м'язи за допомогою горизонтальної бічної сполучнотканинної перегородки поділяються на спинний та черевний відділи (рис. 186). Кожен міомер відокремлений від сусідньої вертикальної сполучнотканинної перегородкою (myoseptum); ці перегородки розташовуються поперечно тулубу. Вертикальні перегородки в ділянці бокової горизонтальної сполучнотканинної перегородки згинаються і утворюють кут, звернений вперед. Верхні (дорсальні), нижні (вентральні) кінці міомерів також згинаються вперед. Виходить ламана лінія між міотомами подібно до такої фігури, де вперед звернений кут, що знаходиться на бічній сполучнотканинній борозні. У вигляді таких конусів і побудовано всю дорсальну мускулатуру нижчих хребетних. Але вже в деяких риб відзначається диференціювання у положенні та напрямку міотомів. У бічній стінці живота частина міотомів розташовується косо до сагітальної площини на різній глибині. Це показує, що у водних тварин з'являються перші ознаки перебудови міотомів, які у наземних хребетних перетворюються на бічні косі м'язи живота. Посередині живота частина міотомів і перетворюється на прямий м'яз живота.
Спинна мускулатура наземних хребетних пристосована до зміщення окремих сегментів тіла, оскільки міотоми перекидаються через ті сполуки, де відбувається рух. У амфібій м'язи спини побудовані ще з самостійних міотомів і нагадують м'язи нижчих хребетних. Тільки у рептилій відзначається зникнення горизонтальної сполучнотканинної перегородки і настає стирання кордону між дорсальною та вентральною мускулатурою. У зв'язку з підвищенням рухливості хребта та грудної клітки частина міотомів зливається у більші м'язи. Так з'являються mm. interspinales, intertransversales, transversospinales, longissimus, iliocostalis і потилично-хребетна група. Це призвело до того, що у вищих хребетних, у тому числі й у людини, первинні міотоми залишилися лише у вигляді коротких м'язів мм. rotatores, що зв'язують послідовно сегменти тіла (хребці).
186. Поверхнева черевна мускулатура тритону (по Maurer).
1 - поджаберний м'яз: 2 - міжм'язова перегородка; 3 – m. obliquus externus superficialis; 4 – m. rectus interims; 5 – m. rectus superficialis.
Черевна мускулатура у нижчих амфібій ще зберігає розподіл як окремих міотомів (рис. 186), але посередині розташовуються поздовжні міотоми, зрощені в прямий м'яз живота. У бічних стінках міотоми змінюють напрямок, але ще утворюють самостійних м'язових шарів. У вищих хребетних міосепти зникають і міотоми зливаються один з одним у великі м'язові пласти, розташовані у три шари. Ці м'язові верстви в людини представлені трьома бічними м'язами. Тільки у прямий м'яз живота у всіх тварин зберігається первинна сегментація у вигляді сухожильних перемичок intersectiones tendineae.
У грудному відділіпо сполучнотканинних перегородках між міотомами проростають ребра, а в міжребер'ях розташовуються міжреберні м'язи, що представляють продовження черевних міотомів. З черевного боку сумчастих m. pyramidalis досягає грудини. У ссавців цей м'яз зберігається у вигляді рудименту.
Філогенез вісцеральної мускулатури. У нижчих хребетних передній відділ кишечника утворений кільцевими волокнами, що охоплюють весь вісцеральний апарат як загального стискача. Починаючи з круглоротих і селяхій, глибокі пучки м'яза вступають у зв'язок з вісцеральними дугами, які прикриті зовні бічними м'язами, пробитими зябровими отворами. У м'язовій масі спостерігається відокремлення окремих м'язів: до верхнього відділу I зябрової дуги прикріплюється м'яз, що піднімає піднебінно-квадратний хрящ (іннервується V парою черепних нервів), до нижнього відділу - м'яз, що призводить до нижньої частини II зябрової дуги (іннервується V парою череп. міжщелепний м'яз, що лежить між гілками нижньої щелепи (іннервується V та VII парами черепно-мозкових нервів). З боку спини позаду зябрових щілин задня частиназагального стискача відокремлюється у m. trapezius.
Починаючи з земноводних, у зв'язку з фіксацією верхньої щелепи до черепа м'язи вісцерального апарату перетворюються. М'яз, що піднімає піднебінно-квадратний хрящ, у амфібій перетворюється на м'яз, що піднімає око, що у вищих тварин редукується. З м'яза, що приводить нижню щелепу, формуються жувальні м'язи.
Міжщелепний м'яз у амфібій і рептилій зберігає своє становище, а у вищих тварин перетворюється на підборідно-під'язичний м'яз. Від міжщелепного м'яза відокремлюється пучок на формування переднього черевця двочеревного м'яза. Всі ці м'язи іннервуються V парою черепно-мозкових нервів.
У водних тварин є м'язи, які іннервуються VII черепним нервом. Сюди відноситься м'яз, що опускає нижню щелепу, яка у ссавців перетворюється на заднє черевце двочеревного м'яза (іннервується VII парою черепних нервів).
У рептилій значного розвитку досягає стискувач шиї (іннервується VII черепним нервом), який у ссавців поділяється на поверхневу та глибоку частини. З глибокої частини розвиваються м'язи навколоротової щілини, з поверхневої - формується решта мімічної мускулатури (іннервується VII парою черепних нервів).
М'язи, пов'язані з зябровим апаратом, зі втратою зябрового типу дихання перетворюються на м'язи гортані, глотки і під'язикові. Трапецієподібний м'яз втрачає зв'язок із зябровими дугами і переходить на плечовий пояс. Від її переднього краю відщеплюється грудино-ключично-соскоподібний м'яз.
У риб та наземних хребетних під'язикові м'язи розвиваються з черевних відростків потиличних міотомів, які огинають ззаду зябровий апарат і на вентральній стороні розташовуються під вісцеральним апаратом. З прямого м'яза живота у земноводних та інших більш високоорганізованих тварин розвиваються м'язи язика, під'язична та м'язи, що лежать нижче під'язичної кістки.
ЛЕКЦІЯ МІОЛОГІЯ ФІЛОГЕНЕЗ, ОНТОГЕНЕЗ І ФУНКЦІОНАЛЬНА АНАТОМІЯ М'язової системи Виконали: Володимирова Я. Б. Кокорєва Т. В.

 М'язи або м'язи (від лат. musculus - мишка, маленька миша) - органи тіла тварин і людини, що складаються з пружної, еластичної м'язової тканини, здатної скорочуватися під впливом нервових імпульсів. Призначені до виконання різних процесів: руху тіла, скорочення голосових зв'язок, дихання. М'язи складаються на 86, 3% із води. У тілі людини 640 м'язів
М'язи або м'язи (від лат. musculus - мишка, маленька миша) - органи тіла тварин і людини, що складаються з пружної, еластичної м'язової тканини, здатної скорочуватися під впливом нервових імпульсів. Призначені до виконання різних процесів: руху тіла, скорочення голосових зв'язок, дихання. М'язи складаються на 86, 3% із води. У тілі людини 640 м'язів
 Мотивація: - - - можливості скоєного руху, обсяг руху; активні чи пасивні рухи запускаються у дію тій чи іншій м'язової групою; впливаючи на м'язовий апарат, ми змінюємо загальний стан; м'язовий рельєф є орієнтиром для топографії судин та нервів; пересадка м'язів, тобто м'яз може переучуватися.
Мотивація: - - - можливості скоєного руху, обсяг руху; активні чи пасивні рухи запускаються у дію тій чи іншій м'язової групою; впливаючи на м'язовий апарат, ми змінюємо загальний стан; м'язовий рельєф є орієнтиром для топографії судин та нервів; пересадка м'язів, тобто м'яз може переучуватися.
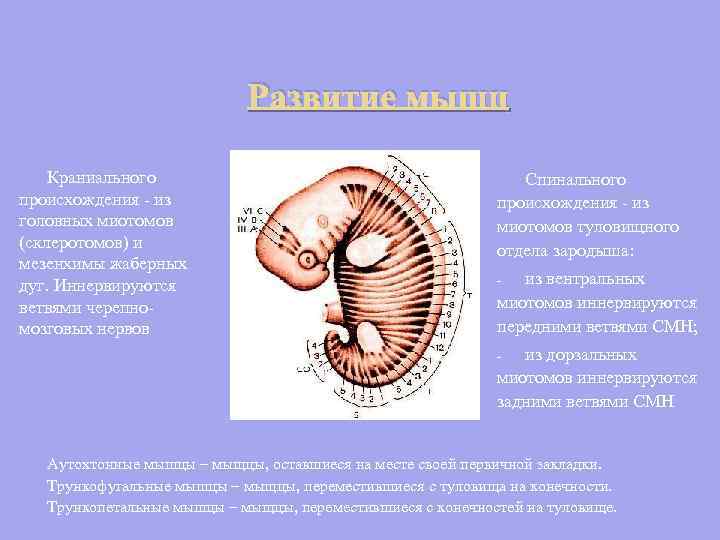 Розвиток м'язів Краніального походження - з головних міотомів (склеротомів) та мезенхіми зябрових дуг. Спинального походження - з міотомів тулубового відділу зародка: з вентральних міотомів іннервуються передніми гілками СМН; - з дорзальних міотомів іннервуються задніми гілками СМН - Аутохтонні м'язи - м'язи, що залишилися на місці своєї первинної закладки. Трункофугальні м'язи - м'язи, що перемістилися з тулуба на кінцівки. Трункопетальні м'язи - м'язи, що перемістилися з кінцівок на тулуб.
Розвиток м'язів Краніального походження - з головних міотомів (склеротомів) та мезенхіми зябрових дуг. Спинального походження - з міотомів тулубового відділу зародка: з вентральних міотомів іннервуються передніми гілками СМН; - з дорзальних міотомів іннервуються задніми гілками СМН - Аутохтонні м'язи - м'язи, що залишилися на місці своєї первинної закладки. Трункофугальні м'язи - м'язи, що перемістилися з тулуба на кінцівки. Трункопетальні м'язи - м'язи, що перемістилися з кінцівок на тулуб.

 Поперечно-смугаста Гладка 1. Одиницею організації є міоцит. Довжина близько 50 мкм. Ширина від 6 мкм. 2. Мимовільне скорочення Контроль з боку автономної нервової системи Рух хвилеподібний, працює повільно, оскільки нервове волокно підходить НЕ до кожної клітини Повільно вступають у дію, але зберігаються на тривалому протязі Не має точної просторової орієнтації клітин 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Серцева Одиницею організації є м'язове волокно-множина міобластів, що плавають у загальній цитоплазмі (саркоплазмі). У них загальна сарколема. Довжина близько 40-100 мм. Ширина від 7 мм. Довільне скорочення Контроль з боку соматичної нервової системи Швидке скорочення, швидка реакція, так кожне м'язове волокно має нервово-м'язовий синапс Швидко включаються, але має короткочасну дію Чітка орієнтація м'язових волокон
Поперечно-смугаста Гладка 1. Одиницею організації є міоцит. Довжина близько 50 мкм. Ширина від 6 мкм. 2. Мимовільне скорочення Контроль з боку автономної нервової системи Рух хвилеподібний, працює повільно, оскільки нервове волокно підходить НЕ до кожної клітини Повільно вступають у дію, але зберігаються на тривалому протязі Не має точної просторової орієнтації клітин 3. 4. 5. 6. 1 2. 3. 4. 5. 6. Серцева Одиницею організації є м'язове волокно-множина міобластів, що плавають у загальній цитоплазмі (саркоплазмі). У них загальна сарколема. Довжина близько 40-100 мм. Ширина від 7 мм. Довільне скорочення Контроль з боку соматичної нервової системи Швидке скорочення, швидка реакція, так кожне м'язове волокно має нервово-м'язовий синапс Швидко включаються, але має короткочасну дію Чітка орієнтація м'язових волокон
 Між м'язовими волокнами знаходяться тонкі прошарки пухкої волокнистої сполучної тканини-ендомізій. Колагенові волокна зовнішнього листка базальної мембрани вплітаються в нього, що сприяє поєднанню зусиль при скороченні міосимпластів. Товстіші прошарки пухкої сполучної тканини оточують по кілька м'язових волокон, утворюючи перимизій і поділяючи м'яз на пучки. Декілька пучків об'єднуються в більш великі групирозділені більш товстими сполучнотканинними прошарками. Сполучну тканину, що оточує поверхню м'яза, називають епімізією.
Між м'язовими волокнами знаходяться тонкі прошарки пухкої волокнистої сполучної тканини-ендомізій. Колагенові волокна зовнішнього листка базальної мембрани вплітаються в нього, що сприяє поєднанню зусиль при скороченні міосимпластів. Товстіші прошарки пухкої сполучної тканини оточують по кілька м'язових волокон, утворюючи перимизій і поділяючи м'яз на пучки. Декілька пучків об'єднуються в більш великі групирозділені більш товстими сполучнотканинними прошарками. Сполучну тканину, що оточує поверхню м'яза, називають епімізією.
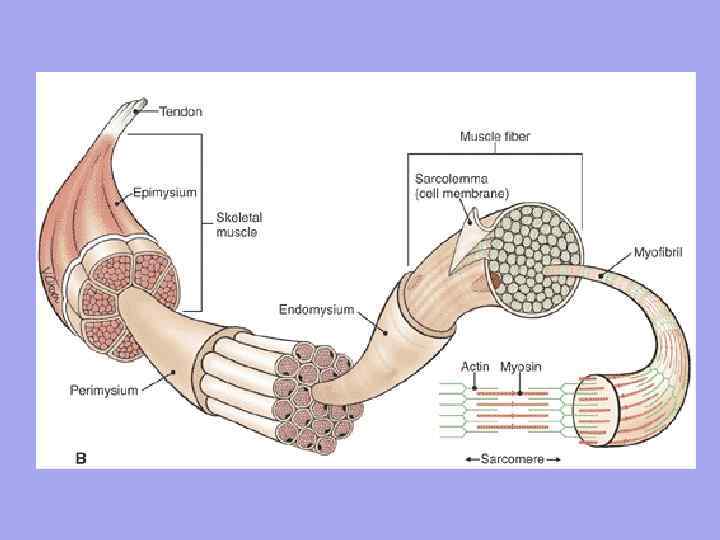
 У м'язі як органі є сполучна тканина Ендомізій – тонка сполучна тканина, що оточує кожне м'язове волокно та невеликі групи волокон. Перімізій – покриває більші комплекси м'язових волокон та м'язові пучки.
У м'язі як органі є сполучна тканина Ендомізій – тонка сполучна тканина, що оточує кожне м'язове волокно та невеликі групи волокон. Перімізій – покриває більші комплекси м'язових волокон та м'язові пучки.
 Значення ендомізія та перимизія 1. Через ендомізіум та перимизіум до м'язового волокна підходять судини та нерви. Утворюють строму органу; 2. М'язові волокна формуються у пучки, пучки у м'яз; 3. Оскільки ендомізій зрощений з сарколеммою м'язового волокна, отже, м'язове волокно, що скорочує, може розтягуватися лише до певної межі.
Значення ендомізія та перимизія 1. Через ендомізіум та перимизіум до м'язового волокна підходять судини та нерви. Утворюють строму органу; 2. М'язові волокна формуються у пучки, пучки у м'яз; 3. Оскільки ендомізій зрощений з сарколеммою м'язового волокна, отже, м'язове волокно, що скорочує, може розтягуватися лише до певної межі.
 Міофібрили у волокні оточені оболонкою – сарколеммою, і занурені у спеціальне середовище – саркоплазму. Залежно від вмісту пігменту та кисню волокна поділяються на білі та червоні. Білі волокна - анаероби, містять більше міофібрил, менше саркоплазми. Запускаються швидко, але довго працювати не можуть. Приклад: грудинно-ключичнососцеподібні, литкові м'язи. Червоні волокна – товсті волокна. Багато міоглобіну в саркоплазмі та цитохрому в мітохондріях, але менше міофібрил. Повільно запускаються, але працюють довго. Приклад: м'яз спини, діафрагма.
Міофібрили у волокні оточені оболонкою – сарколеммою, і занурені у спеціальне середовище – саркоплазму. Залежно від вмісту пігменту та кисню волокна поділяються на білі та червоні. Білі волокна - анаероби, містять більше міофібрил, менше саркоплазми. Запускаються швидко, але довго працювати не можуть. Приклад: грудинно-ключичнососцеподібні, литкові м'язи. Червоні волокна – товсті волокна. Багато міоглобіну в саркоплазмі та цитохрому в мітохондріях, але менше міофібрил. Повільно запускаються, але працюють довго. Приклад: м'яз спини, діафрагма.
 Кожен м'яз має мережу кровоносних судин. Скорочення м'яза сприяє току крові. У розслабленому непрацюючому м'язі більшість кровоносних капілярів закрита для струму крові. При скороченні м'язів всі кровоносні капіляри відразу відкриваються.
Кожен м'яз має мережу кровоносних судин. Скорочення м'яза сприяє току крові. У розслабленому непрацюючому м'язі більшість кровоносних капілярів закрита для струму крові. При скороченні м'язів всі кровоносні капіляри відразу відкриваються.

 Будова м'яза Кожен м'яз одним кінцем з'єднується з однією кісткою (початок м'яза), а іншим – з іншого (прикріплення м'яза). У м'язі розрізняють: голівку черевця хвостову частину.
Будова м'яза Кожен м'яз одним кінцем з'єднується з однією кісткою (початок м'яза), а іншим – з іншого (прикріплення м'яза). У м'язі розрізняють: голівку черевця хвостову частину.
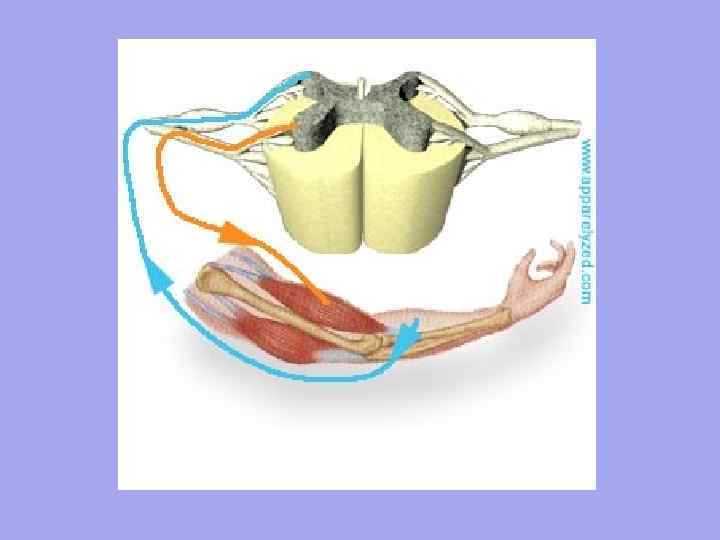
 До кожного м'язового волокна підходять рухові нервові волокна і відходять чутливі нервові волокна. Кількість нервових закінчень у м'язі залежить від ступеня функціональної активності м'язів.
До кожного м'язового волокна підходять рухові нервові волокна і відходять чутливі нервові волокна. Кількість нервових закінчень у м'язі залежить від ступеня функціональної активності м'язів.
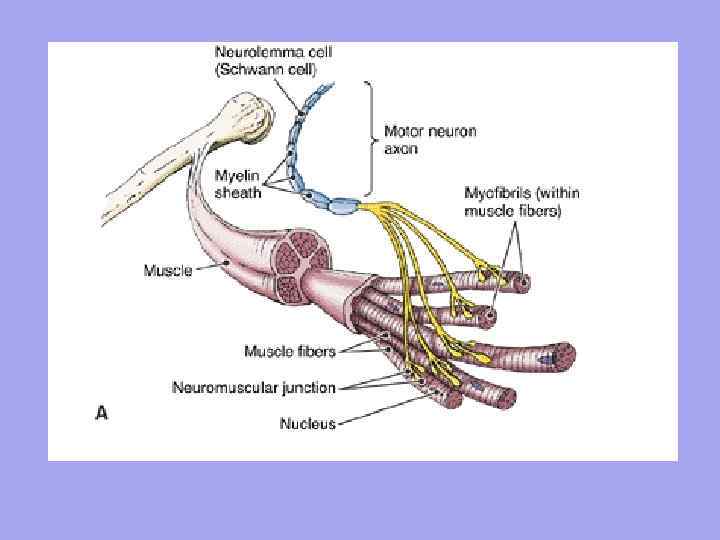
 Кожне м'язове волокно іннервується самостійно і оточене мережею гемокапілярів, утворюючи комплекс, що називається міоном. Група ж м'язових волокон, що іннервуються одним мотонейроном, називається моторною одиницею. Характерно, що м'язові волокна, що належать до однієї моторної одиниці, лежать не поряд, а розташовані мозаїчно серед волокон, що належать до інших одиниць.
Кожне м'язове волокно іннервується самостійно і оточене мережею гемокапілярів, утворюючи комплекс, що називається міоном. Група ж м'язових волокон, що іннервуються одним мотонейроном, називається моторною одиницею. Характерно, що м'язові волокна, що належать до однієї моторної одиниці, лежать не поряд, а розташовані мозаїчно серед волокон, що належать до інших одиниць.

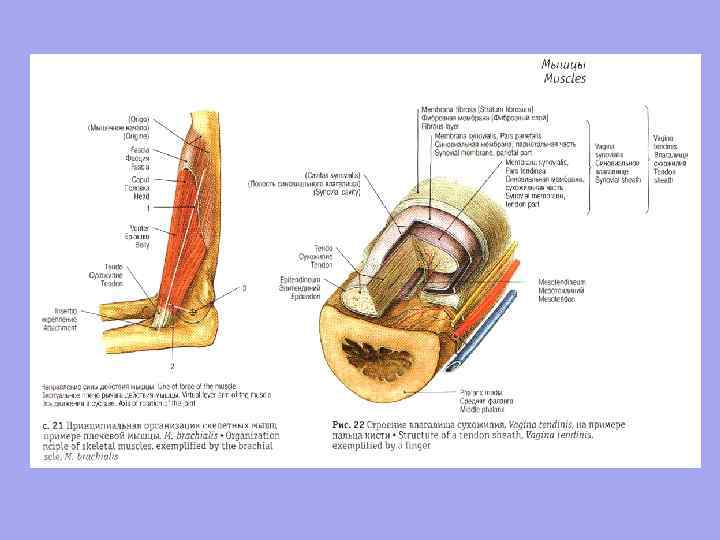
 Сухожилля – це щільний волокнистий сполучно-тканинний тяж, яким м'яз починається або прикріплюється до скелета.
Сухожилля – це щільний волокнистий сполучно-тканинний тяж, яким м'яз починається або прикріплюється до скелета.
 перитеноніум колагенові волокна IV типу ендотеноніум Колагенові волокна сухожилля, що переплітаються з колагеновими волокнами окістя, вплітаються в основну речовину кісткової тканини, утворюючи на кістках гребені, горбики, бугристості, западини, вдавлення.
перитеноніум колагенові волокна IV типу ендотеноніум Колагенові волокна сухожилля, що переплітаються з колагеновими волокнами окістя, вплітаються в основну речовину кісткової тканини, утворюючи на кістках гребені, горбики, бугристості, западини, вдавлення.
 Фасції – це сполучнотканинні колагенні волокна з невеликою домішкою еластичних волокон Поверхнева скронева фасція Глибокі фасції стегна
Фасції – це сполучнотканинні колагенні волокна з невеликою домішкою еластичних волокон Поверхнева скронева фасція Глибокі фасції стегна
 1. 2. 3. 4. 5. Фасції відокремлюють м'язи від шкіри і усувають зміщення шкіри при рухах м'язів, що скорочуються. Фасції економлять силу скорочення м'язів, усуваючи тертя між м'язами під час скорочення. Фасції розтягують великі вени при натягу, внаслідок чого кров з периферії «присмоктується» у ці вени. Фасції мають значення як бар'єри, що перешкоджають поширенню інфекції та пухлин. Під час операцій фасції допомагають визначати розташування м'язів, кровоносних судин, нутрощів.
1. 2. 3. 4. 5. Фасції відокремлюють м'язи від шкіри і усувають зміщення шкіри при рухах м'язів, що скорочуються. Фасції економлять силу скорочення м'язів, усуваючи тертя між м'язами під час скорочення. Фасції розтягують великі вени при натягу, внаслідок чого кров з периферії «присмоктується» у ці вени. Фасції мають значення як бар'єри, що перешкоджають поширенню інфекції та пухлин. Під час операцій фасції допомагають визначати розташування м'язів, кровоносних судин, нутрощів.
 Класифікація м'язів Скелетні м'язи різноманітні за формою, структурою, положенням щодо осей суглобів і т. д., тому класифікуються по-різному.
Класифікація м'язів Скелетні м'язи різноманітні за формою, структурою, положенням щодо осей суглобів і т. д., тому класифікуються по-різному.

 ІІІ. По функціональній особливості Статичні (сильні) – коротке черевце та довге сухожилля. М'язи працюють із більшою силою, але з меншим розмахом руху. Динамічні (спритні) - довгі м'язові пучки, сухожилля короткі. М'язи працюють з меншою силою, але роблять рухи більшого розмаху
ІІІ. По функціональній особливості Статичні (сильні) – коротке черевце та довге сухожилля. М'язи працюють із більшою силою, але з меншим розмахом руху. Динамічні (спритні) - довгі м'язові пучки, сухожилля короткі. М'язи працюють з меншою силою, але роблять рухи більшого розмаху
 Допоміжний апарат м'язів Скелетні м'язи мають допоміжний апарат, який полегшує їхнє функціонування. n n Фасції; Кістково-фасціальні піхви; синовіальні сумки; Синовіальні піхви сухожиль; М'язові блоки; Сесамоподібні кістки.
Допоміжний апарат м'язів Скелетні м'язи мають допоміжний апарат, який полегшує їхнє функціонування. n n Фасції; Кістково-фасціальні піхви; синовіальні сумки; Синовіальні піхви сухожиль; М'язові блоки; Сесамоподібні кістки.
 Аномалії розвитку м'язів Зустрічаються дуже часто і поділяються на три групи: 1. Відсутність будь-якого м'яза; 2. Наявність додаткового м'яза, якого немає в природі. 3. Додаткові пучки наявного м'яза.
Аномалії розвитку м'язів Зустрічаються дуже часто і поділяються на три групи: 1. Відсутність будь-якого м'яза; 2. Наявність додаткового м'яза, якого немає в природі. 3. Додаткові пучки наявного м'яза.
 Недорозвиток грудинно-ключично-соскоподібного м'яза - Кривошия Недорозвиток діафрагми. Причина - поява діафрагмальних гриж. Недорозвиток дельтоподібного та трапецієподібного м'язів – Деформація плечового пояса та плеча
Недорозвиток грудинно-ключично-соскоподібного м'яза - Кривошия Недорозвиток діафрагми. Причина - поява діафрагмальних гриж. Недорозвиток дельтоподібного та трапецієподібного м'язів – Деформація плечового пояса та плеча


 I. За формою: Веретеноподібні; Стрічкоподібні; Плоскі широкі; Зубчасті; Довгі; n n n Квадратні; Трикутні; Круглі; Дельтоподібна; Камбалоподібна і т.д.
I. За формою: Веретеноподібні; Стрічкоподібні; Плоскі широкі; Зубчасті; Довгі; n n n Квадратні; Трикутні; Круглі; Дельтоподібна; Камбалоподібна і т.д.



 ІІ. У напрямку м'язових волокон З прямими паралельними волокнами; З поперечними; З круговими; Перисті: A. Одноперисті; Двоперисті; C. Багатоперисті. B.
ІІ. У напрямку м'язових волокон З прямими паралельними волокнами; З поперечними; З круговими; Перисті: A. Одноперисті; Двоперисті; C. Багатоперисті. B.

 IV. За функцією: Привідні; Відвідні; Згинальні; Розгинальні; Пронатори; n Супінатори; Напружуючі; М'язи – синергісти; М'язи – антагоністи.
IV. За функцією: Привідні; Відвідні; Згинальні; Розгинальні; Пронатори; n Супінатори; Напружуючі; М'язи – синергісти; М'язи – антагоністи.
 V. Стосовно суглоба: Односуглобові; Двосуглобові; Багатосуглобові.
V. Стосовно суглоба: Односуглобові; Двосуглобові; Багатосуглобові.
Чи не ізольована м'язова система
Єдиний шкірно-м'язовий мішок
Поява поперечно-смугастої м'язової тканини
Поділ м'язових тяжів на міотоми.
Розвиток груп м'язів
Розвиток м'язів кінцівок (зміна довкілля)
Розвиток діафрагми
Розвиток усіх груп м'язів – виконання диференційованих рухів
2 Онтогенез м'язової системи: джерела та терміни розвитку.
Похідні міотома: з дорзальної ділянки розвиваються м'язи спини
з вентрального - м'язи грудей та живота
Мезенхіма - м'язи кінцівок
I вісцеральна дуга (ВД) - жувальні м'язи
II ВД – мімічні м'язи
III та IV ВД – м'язи м'якого піднебіння, глотки, гортані, верхньої частини стравоходу
V ВД - грудино-ключично-соскоподібні та трапецієподібні м'язи
З потиличних міотомів – м'язи язика
З міотомів - м'язи очного яблука
М'язи розвиваються з мезодерми. На тулуб вони виникають із первинної сегментованої мезодерми – сомітів: 3-5 потиличних, 8 шийних, 12 грудних, 5 поперекових, 5 крижових, 4-5 куприкових.
Кожен соміт поділяється на склеротом, дерматом та міотом- З нього і розвиваються м'язи тулуба. Сомити з'являються рано, коли довжина зародка становить 10-15 мм.
З дорсальнихчастин міотомів виникають глибокі, власні(аутохтонні) м'язи спини, вентральних- Глибокі м'язи грудей і живота. Вони закладаються, розвиваються та залишаються в межах тулуба – тому називаються аутохтонними (місцевими, тубільними). Дуже рано міотоми зв'язуються з нервовою системоюта кожному м'язовому сегменту відповідає нервовий сегмент. Кожен нерв слідує за м'язом, що розвивається, вростає в неї і, поки вона не диференціювалася, підпорядковує своєму впливу.
У процесі розвитку частина скелетних м'язів переміщається з тулуба та шиї на кінцівки. трункофугальнім'язи: трапецієподібні, грудино-ключично-соскоподібні, ромбоподібні, що піднімає лопатку та ін. Частина м'язів, навпаки, прямує з кінцівок на тулуб – гробкопетальнім'язи: широка м'яз спини, велика і мала грудні, велика поперекова.
М'язи головимімічні та жувальні, над- та під'язикові м'язи шиї розвиваються з несегментованої вентральної мезодерми, що входить до складу вісцеральних (жаберних) дуг. Вони називаються вісцеральними і, наприклад, жувальні м'язи розвиваються на основі першої вісцеральної дуги, а мімічні – другий. Однак м'язи очного яблука та язика розвиваються із потиличних міотомів сегментованої мезодерми. Глибокі передні та задні м'язи шиї також виникають із потиличних шийних міотомів, а поверхнева та середня групам'язів у передній ділянці шиї розвиваються на основі несегментованої мезодерми вісцеральних дуг.
3 М'яз: визначення, будова.
Мускул(м'яз) – орган, побудований з м'язових волокон (клітин), кожне з них має сполучно-тканинну оболонку – ендомізієм. У пучки м'язові волокна поєднує інша фіброзна оболонка - перимизій, а весь м'яз полягає в загальну фіброзну оболонку, утворену фасцією - епімізій. Між пучками проходять судини та нерви, що забезпечують м'язові волокна.
На макрорівні скелетний м'яз має:
· черевце(вентер) - м'ясиста частина органу, що займає його середину;
· сухожилля(Тендо), що відноситься до дистального кінця, воно може бути у вигляді апоневрозу, сухожильних перемичок, довгих пучків поздовжніх фіброзних волокон;
· головку, Що становить проксимальну частину;
· Сухожилля та головка кріпляться на протилежних кінцях кісток.
Проксимальне сухожилля або головка м'яза - початок м'яза на кістці знаходиться ближче до серединної осі тіла - фіксована точка (punctum fixum) (як правило збігається з початком м'яза). Дистальне сухожилля, «хвіст» – кінець м'яза лежить на кістці дистальніше і, будучи місцем прикріплення, називається рухомою точкою (punctom mobile). При скороченні м'язи точки зближуються, а зміні положення тіла можуть змінюватися місцями.
Сухожилля різні формою: тонкі довгі сухожилля мають м'язи кінцівок; м'язи, що у формуванні стінок черевної порожнини, мають широке плоске сухожилля, розташоване між двома черевками - сухожильне розтягування чи апоневроз.
4 Класифікація м'язів за походженням, будовою, формою та функцією.
А. Везалій – анатом епохи Відродження – позначав м'язи цифрами, але зараз вони класифікуються за іншими принципами.
За походженням:
- з дорсальнихчастин міотомів виникають глибокі, власні(аутохтонні) м'язи спини
- з вентральних- Глибокі м'язи грудей і живота, вони закладаються, розвиваються і залишаються в межах тулуба - тому називаються аутохтонними (місцевими, тубільними).
За функцією розрізняють:
· М'язи- антагоністи, як то: згиначі та розгиначі, що приводять і відводять, супінатори та пронатори – такі м'язи діють у протилежних напрямках;
· М'язи- синергісти– діють за одним напрямом, посилюючи один одного; при ряді рухів як синергісти виступають і м'язи-антагоністи, наприклад, при виконанні кругових рухів;
· Головні та допоміжні м'язи.
За розташуванням:
· Зовнішні та внутрішні
· Поверхневі та глибокі
· Медіальні та латеральні
За формою та будовою:
· Веретеноподібні м'язи (musculi fusiformes) - явл. довгими важелями (двоголовий м'яз плеча)
· широкі м'язи - беруть участь в утворенні стінок тулуба (прямий м'яз живота)
· Одно-, дво- та багатоперисті м'язи – залежно від того, по один бік від сухожилля або на дві і більше сторін лягають м'язові пучки, наприклад, багатоперистий дельтовидний м'яз.
· м'язи, форма яких відповідає певному геометричній фігурі, наприклад, ромбовидна велика і мала, трапецієподібна, квадратна, кругова, пряма, тонка;
· М'язи, що мають кілька головок або черевців: дво-, три-, чотириголові м'язи кінцівок; двочеревна на шиї;
· М'язи, в назвах яких відображено напрямок волокон: поперечна, поздовжня, коса;
· М'язи, в назвах яких відображена функція: розгинач, згинач, що приводить, відводить, що піднімає, опускає, стискає і т. д.;
· М'язи, великі за площею та довжиною: широкі та найширші, великі та малі, довгі та короткі;
· Одно-, дво- та багатосуглобові м'язи залежно від того, на скільки суглобів діють м'язи, є м'язи які взагалі не діють на суглоб.
Також розрізняють:
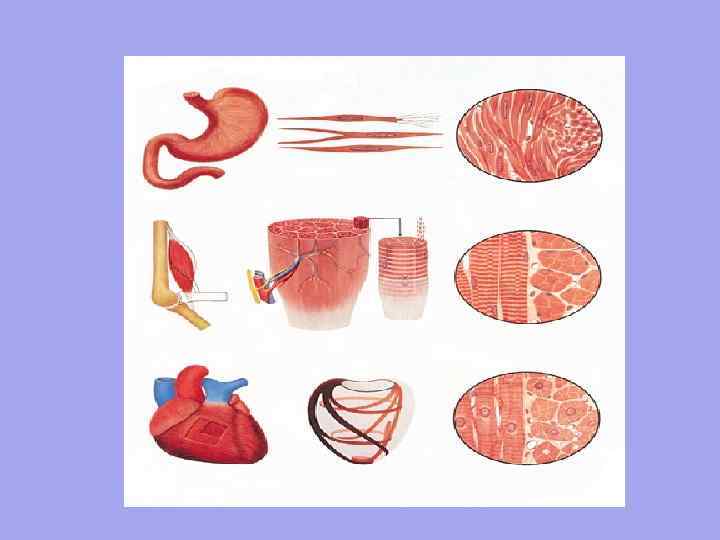
- гладкі м'язи (мимовільна мускулатура) - розвиваються з вісцерального листка сплахнотома, розташовуються в стінці внутрішніх органів, скорочуються мимовільно, іннервуються вегетативною нервовою системою;
- поперечно-смугасті м'язи – скелетні (довільна муск., розв. з міотомів – обр. скелет. мускулатуру, инерв.- сомат.нерв.сист) і серцева (мимовільна мускулатура, має попер-полос строй., але склад. з відділ. клітин – кардіоміоцитів, інерв. – вегет.нерв.сист.)
Скелетні м'язи виконують долає, поступається роботущо забезпечує м'язову динаміку тіла, утримуючу- Міостатичну роботу.
Соматична та вісцеральна м'язова система, її філо-онтогенез. Підшкірні м'язи. Скелетні м'язи. Будова м'яза, як органу. Класифікація м'язів Допоміжні пристрої м'язів.
Міологія(Myologiа) - це розділ анатомії свійських тварин, що вивчає будову м'язової системи. М'язова тканина, що є основою цієї системи, здійснює всі рухові процеси в організмі тварин. Завдяки їй тіло фіксується у певному положенні та переміщається у просторі, здійснюються дихальні рухи грудної клітки та діафрагми, рух очей, ковтання, рухові функції внутрішніх органів, включаючи роботу серця.
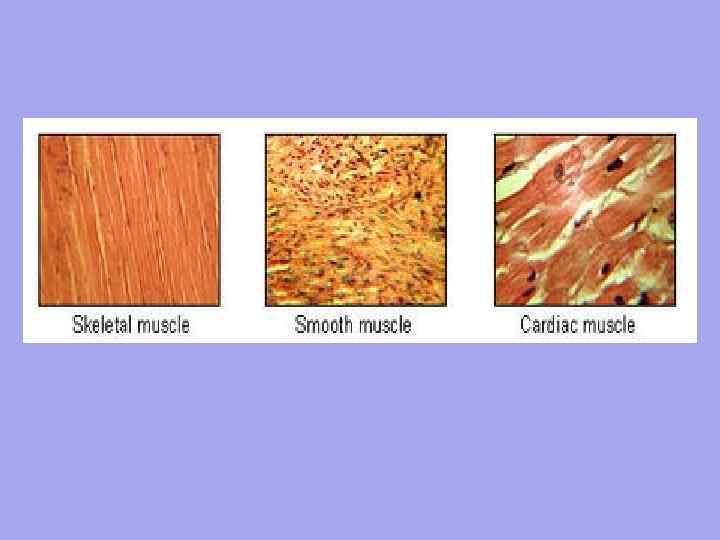
М'язова тканинамає спеціальні скорочувальні органели - міофібрили . Міофібрили,що складаються з тонких білкових ниток (міофіламентів), можуть бути невичерпаними або смугастими (поперечно-смугастими). Відповідно розрізняють неисчерченную і исчерченную м'язову тканину.
1) Невичерпана м'язова тканина складається з клітин (гладких міоцитів) веретеноподібної форми. Ці клітини утворюють м'язові шари у стінках кровоносних та лімфатичних судин, у стінках внутрішніх органів (шлунок, кишечник, сечовивідні шляхи, матка тощо). Довжина клітин коливається від 20 мкм (у стінці кровоносної судини) до 500 мкм (у стінці матки тільної корови), діаметр від 2 до 20 мкм. У функціональному відношенні невичерпана м'язова тканина має ряд особливостей: вона має велику силу (наприклад, у кишечнику постійно пересуваються значні маси їжі), має слабку стомлюваність, повільне скорочення і ритмічність рухів (у стінці кишечника невикреслена м'язова тканина скорочується 12 разів на хвилину, селезінці - лише 1 раз).
2) Смугаста м'язова тканина характеризується наявністю смугастих міофібрил, має 2 різновиди.
А) Смугаста серцева м'язова тканина складається з подовжених клітин (кардіоміоцитів) квадратної форми. Їхні кінці, з'єднуючись один з одним у ланцюжки, формують так звані функціональні м'язові "волокна" товщиною 10-20 мкм. Тісно зв'язуючись між собою, функціональні м'язові "волокна" утворюють м'язову оболонку серця ( міокард), постійні та ритмічні скорочення якого приводять у рух кров.
Б) Исчерченная скелетна м'язова тканина, на відміну серцевої, складається з клітин, та якщо з багатоядерних м'язових утворень (миосимпластов) циліндричної форми. Довжина міосимпластів коливається від кількох міліметрів до 13-15 см, діаметр від 10 до 150 мкм. Кількість ядер у них може сягати кількох десятків тисяч. Міосимпласти (їх ще називають "м'язовими волокнами") утворюють скелетні м'язи і входять до складу деяких органів (мова, ковтка, горло, стравохід та ін.). У функціональному відношенні скелетна м'язова тканина легко збудлива і скорочується швидше, ніж неисчерченная (наприклад, у звичайних умовах скелетний м'яз скорочується протягом 0,1 с, а неисчерченная - протягом кількох секунд). Але, на відміну гладких (неисчерченных) м'язів внутрішніх органів, скелетні м'язи швидше втомлюються.
М'язову системузалежно від особливостей будови, характеру рухової функції та іннервації поділяють на соматичну та вісцеральну.
Соматична м'язова система складає 40% від маси тіла і побудована з міосімпластів. Вона довільна та іннервується соматичною нервовою системою. Соматичні м'язи скорочуються швидко, енергійно, але короткочасно та швидко втомлюються. Такий тип скорочення називається тетанічнимі він уражає соматичної мускулатури. До неї належать:
1) підшкірні м'язи, які не мають зв'язку зі скелетом та прикріплюються до шкіри; їх скорочення викликають посмикування шкіри і дозволяють їй збиратися в дрібні складки;
2) скелетна мускулатура, що закріплюється на скелеті;
3) діафрагма – куполоподібний м'яз, що відокремлює грудну порожнину від черевної;
4) м'язи язика, глотки, гортані, вушної раковини, очного яблука, середнього вуха, стравоходу та зовнішніх органів розмноження.
Вісцеральна м'язова система становить 8% від маси тіла та побудована з гладких міоцитів. Вона мимовільна та іннервується вегетативною нервовою системою. Гладкі м'язи скорочуються повільно, тривалий час і не вимагають великої кількості енергії. Такий тип скорочення називають тонічним тавін характерний для вісцеральної мускулатури, що утворює м'язові пучки, шари та оболонки внутрішніх органів.
Філо-онтогенез м'язової системи
У філогенезі хордових м'язова система послідовно проходить ряд стадій.
У ланцетникавона представлена парним поздовжнім м'язом (правим і лівим), що йде вздовж тіла і розділяється сполучнотканинними перегородками (міосептами) на короткі прямі м'язові пучки (міомери). Такий (сегментарний) поділ єдиного м'язового пласта називається метамерією.
Зі збільшенням рухливості, відокремленням голови та розвитком кінцівки (у вигляді плавців) у рибвідбувається поділ поздовжнього м'яза горизонтальною септою на дорсальний і вентральний м'язи, а також
Відокремлення мускулатури голови, тулуба, хвоста та плавників.
З виходом на сушу та збільшенням різноманітності рухів у амфібій та рептилійвідбувається поділ дорсального м'яза, так само як і вентрального, на два тяжи: латеральний (поперечно-реберний м'яз) і медіальний (поперечно-остистий м.). Крім цього, у рептилій із латерального тяжу вперше з'являються підшкірні м'язи, які прикріплюються до шкіри.
У більш високоорганізованих тварин ( птахів та ссавців) відбувається подальша диференціація м'язової системи: латеральний та медіальний тяжі, кожен з них, поділяються на два шари (поверхневий та глибокий). Крім цього, у ссавців уперше з'являється діафрагма.
Філогенез м'язової системи.
| Хордові | М'язова система | |||||||
| Ланцетник | Поздовжній м'яз | |||||||
| Риби | Дорсальна | Вентральна | ||||||
| Амфібії, рептилії | Латеральна | Медіальна | Латеральна | Медіальна | ||||
| Птахи, ссавці | Повір | Глибока. | П | Г | П | Г | П | Г |
В онтогенезі м'язова система в основному розвивається з міотомів мезодерми, виняток становлять деякі м'язи голови та шиї, які утворюються з мезенхіми (трапецієподібна, плечоголовна).
На початку формується м'язовий поздовжній тяж, який відразу диференціюється на дорсальний і вентральний пласти; Далі кожен з них поділяється на латеральний та медіальний пласти, які, у свою чергу, диференціюються на поверхневий та глибокий шари, останні дають початок певним групам м'язів. Наприклад, з поверхневого шару латерального пласта розвиваються здухвинно-реберні м'язи, та якщо з глибокого шару латерального пласта – найдовші м'язи спини, шиї, голови.
Підшкірні м'язи – musculi cutanei
Підшкірні м'язи прикріплюються до шкірного покриву, фасцій і немає зв'язку зі скелетом. Їх скорочення викликають підсмикування шкіри і дозволяють їй збиратися в дрібні складки. До цих м'язів належать:
1) Підшкірний м'яз шиї – m. Cutaneus colli (особливо сильно розвинена у собак). Вона йде вздовж шиї, ближче до її вентральної поверхні та переходить на лицьову поверхню до м'язів рота та нижньої губи.
2) Підшкірний м'яз лопатки та плеча (лопатково-плечовий) – m. Cutaneus omobrachialis. Вона покриває область лопатки та частково плече. Добре виражена у коня і великої рогатої худоби.
3) Підшкірний м'яз тулуба – m. Cutaneus trunci. Вона розташована з боків від грудної та черевної стінок і каудально віддає пучки в колінну складку.
4) У самок в області молочних залоз розташовуються краніальні та каудальні м'язи молочної залози (mm. Supramammilaris cranialis et caudalis), які надають складчастості шкірі та допомагають виведенню молока. Сильно розвинені у хижих тварин.
У самців у цій галузі є краніальні та каудальні препуціальні м'язи (mm.preputialis cranialis et caudalis), які забезпечують складчастість препуція та виконують роль його сфінктера.
Скелетна мускулатура
Скелетна мускулатура є активною частиною опорно-рухового апарату. Вона складається з скелетних м'язів та їх допоміжних пристроїв, до яких відносяться фасції, синовіальні сумки, синовіальні піхви сухожиль, блоки, сезамоподібні кістки.
У тілі тварини налічується близько 500 кістякових м'язів. Більшість їх парні і розташовуються симетрично з обох боків тіла тварини. Їхня сумарна маса становить у коня 38-42% від маси тіла, у великої рогатої худоби 42-47%, у свиней 30-35% від маси тіла.
М'язи в тілі тварини розташовуються не безладно, а закономірно в залежності від дії сили тяжкості тварини і роботи, що виконується. Вони впливають ті частини скелета, які пов'язані рухомо, тобто. м'язи діють на суглоби, синдесмози.
Основними місцями прикріплення м'язів є кістки, але іноді вони прикріплюються до хрящів, зв'язків, фасцій, шкіри. Вони покривають скелет так, що кістки лише у деяких місцях лежать безпосередньо під шкірою. Закріплюючись на скелеті, як на системі важелів, м'язи при своєму скороченні викликають різні рухи тіла, фіксують скелет у певному положенні та надають форму тілу тварини
Основні функції скелетних м'язів:
1) Основна функція м'язів – динамічна. Скорочуючись, м'яз коротшає на 20-50% своєї довжини і тим самим змінює положення пов'язаних з нею кісток. Виконується робота, результатом якої є рух.
2) Інша функція м'язів – статична. Виявляється вона у фіксації тіла у певному положенні, у збереженні форми тіла та його частин. Один із проявів цієї функції – здатність спати стоячи (кінь).
3) Участь в обміні речовин та енергії. Скелетні м'язи є «джерелами тепла», оскільки за її скорочення близько 70% енергії перетворюється на тепло і лише 30% енергії забезпечує рух. У скелетних м'язах утримується близько 70% води організму, тому їх ще називають джерелами води. Крім цього, між м'язовими пучками і всередині них може накопичуватися жирова тканина (особливо при відгодівлі свиней).
4) Одночасно, при роботі скелетні м'язи допомагають роботі серця, проштовхуючи венозну кров по судинах. В експериментах вдалося з'ясувати, що скелетні м'язи діють подібно до насоса, забезпечуючи рух крові по венозному руслу. Тому скелетні м'язи ще називають "периферичними м'язовими серцями".
Будова м'язу з погляду біохіміка
Скелетний м'яз складається з органічних та неорганічних сполук. До неорганічних сполук відносяться вода та мінеральні солі (солі кальцію, фосфору, магнію). Органічну речовину переважно представлено білками, вуглеводами (глікоген), ліпідами (фосфатиди, холестерин).
Таблиця 2. Хімічний складскелетного м'яза
Хімічний склад скелетних м'язів схильний до значних вікових і меншою мірою видових, породних і статевих відмінностей, що насамперед пов'язано з неоднаковим вмістом у них води (з віком % води зменшується).


